

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Recent emerging data have suggested that dyslipidemia, characterized by the elevation of low-density proteins, may be associated with cardiovascular disease, which is the primary cause of death in Westernized countries.12 Atherosclerosis is a chronic inflammatory disease principally associated with high plasma concentrations of low-density lipoprotein (LDL). It is a complex progressive disease that involves deposition of plaques within the arteries, which are composed of fat, cholesterol, calcium, and immune cells. The first step in the pathogenesis of atherosclerosis includes endothelial dysfunction, characterized by increased cell permeability for adhesion molecules. Second, lipids accumulate within the artery, which consists of monocyte-derived macrophages and T lymphocytes. Third, a fibrous cap in the lumen of the artery forms due to necrosis, advancing and worsening the lesion. Finally, the unstable fibrous plaque causes thrombosis and occlusions within the artery.3 Atherosclerosis can cause morbidity and/or mortality due to thrombosis and subsequent myocardial infarction or stroke, chronic kidney disease, peripheral artery diseases, and angina.34
Mechanisms involved in the development of atherosclerosis have been intensively studied in animal models with a goal of preventing atherosclerotic lesion development or even alleviating its symptoms.5 According to numerous studies in atherosclerosis mouse models, the immune system plays a major role in atherosclerosis or plaque formation.6 However, this may differ in patients with atherosclerosis. Nonetheless, there is ample evidence that complex immune responses are responsible for the initiation and progression of atherosclerosis. Therefore, the immune system may be a therapeutic target for preventing ischemic damage to tissues during atherosclerosis, which would otherwise cause cardiovascular events and possibly death.
Both innate and adaptive immune responses play a role during the pathogenesis of atherosclerosis. Hypercholesterolemia and the elevation of plasma LDL levels drive the formation and accumulation of lipid-laden atherosclerotic plaques in the artery wall. This is primarily associated with vascular inflammation, while activated macrophages and T helper type (Th) 1 cells promote plaque formation.7 The continuous deposition of LDL results in endothelial dysfunction, followed by the upregulation of adhesion molecules for recruiting circulating monocytes. These macrophages then transform into foam cells, and in turn, secrete interleukin (IL)-1β, while activated Th1 cells secrete interferon (IFN)-γ to exacerbate the inflammatory response.78 The heterogeneity of immune cells and mediators differentially affects the balance between progression and resolution of plaque inflammation.6
Intensive investigation has been carried out on the interdependence of immune cell fate and metabolism since they have been found to be interrelated at the molecular, cellular, organ, and organism level. For example, cases with type 1 and type 2 diabetes have increased risk of developing cardiovascular diseases like atherosclerosis, which might be due to other metabolic disturbances such as hyperglycemia and hyperlipidemia. Furthermore, metabolic syndrome consists of abdominal obesity and hypertension, as well as high blood glucose, high serum triglycerides (TG), low high-density lipoprotein, and high LDL levels. This also increases the risk of developing type 2 diabetes and cardiovascular diseases.9 In this review, we specifically describe the role of the metabolic activation of distinct immune cells in the development of atherosclerosis (Fig. 1).
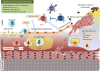 | Fig. 1The stages of atherosclerotic development are as follows. 1) endothelial dysfunction characterized by increased permeability for adhesion molecules, 2) accumulation of lipids within the artery to fatty streaks consisting of monocyte-derived macrophages and T lymphocytes, 3) necrotic core-mediated fibrous cap formation in the lumen of advanced and complex lesions, and 4) unstable fibrous plaque leading to hemorrhage, thrombosis, and occlusions to the artery.oxLDL, oxidized low-density lipoprotein; Ig, immunoglobulin; Ang II, angiotensin II; LDL, low-density lipoprotein; TGF, transforming growth factor; IFN, interferon; MMP, metalloproteinase; Treg, regulatory T cell; Mox, macrophage induced by phospholipids; VSMC, vascular smooth muscle cells.
|
FACTORS IN THE PROGRESSIVE DEVELOPMENT OF ATHEROSCLEROSIS
LDL modification by oxidation, glycation, and aggregation, plays an important role in the initiation of endothelial dysfunction and the activation of immune cells in atherosclerotic cardiovascular disease.3 Oxidized LDL (oxLDL) can elicit an innate inflammatory response, which is further enhanced by myeloperoxidase, lipoxygenase, and reactive oxygen species.10 According to genetic studies, prospective epidemiologic cohort studies, Mendelian randomization studies, and randomized intervention trials by separate meta-analysis, cumulative LDL arterial burden is highly associated with the progression of atherosclerotic cardiovascular diseases. Because of these findings, clearance of LDL and cholesterol by statins (HMG-CoA reductase inhibitor) and alirocumab/evolocumab (proprotein convertase subtilisin/kexin type 9 inhibitor), respectively, have been identified as therapeutic targets.1 Furthermore, the activity of cholesterol acyltransferase and cholesterol ester hydrolase increases during the differentiation of monocytes to macrophages, resulting in foam cells characterized by neutral lipid accumulation.11 These enzymes are encoded by ABCA1 and ABCG1; regulation of these genes that control cholesterol efflux capacity may be a therapeutic target for foam cell-mediated atherosclerosis. Finally, LDL particles in the arterial intima cause differentiation of infiltrating monocytes into macrophages, a major cell population dynamically characterized by its heterogeneity and complexity.12
In addition, cells expressing major histocompatibility complex class II receptors (T cells) can be activated by antigen presenting cells, such as dendritic cells (DCs) displaying plaque-derived antigen and B cells. These cells respond to the epitopes of oxLDL, lysophosphatidylcholine, phosphorylcholine, and apo-lipoprotein B100 in the atherosclerotic plaques.7 Hyperhomocysteinemia promotes atherosclerosis by activating the nucleotide-binding domain-like receptor protein 3 (NLRP3) inflammasome, and it was found that gene silencing of upregulated NLRP3 significantly attenuates atherosclerotic lesion formation that were induced by a high fat and methionine diet.13 Likewise, the Canakinumab Anti-Inflammatory Thrombosis Outcomes Study highlighted that modulation of the immune system could be a therapeutic approach to treat atherosclerosis-related cardiovascular diseases, independent of a lipid-lowering effect.14 Based on unsupervised clustering of a transcriptional landscape for total viable leukocytes (CD45+) from atherosclerotic aortas of Western diet-fed LDL receptor (Ldlr)−/− mice, 13 distinct immune cells types were identified: 3 macrophage subsets, 1 DC, T-cell subsets, 1 B cell, 1 mast cell, 1 monocyte, 1 granulocytes and 1 natural killer cell.15 These cells correlated with different contributions at different lesions and atherosclerosis development time points.15 Metabolic reprogramming mediated by dyslipidemia, including hypercholesterolemia and oxLDL, in individual immune cells has been shown by novel technologies like single-cell RNA sequencing (scRNA-seq) analysis and specialized liquid chromatography tandem mass spectrometry for metabolic flux profiling. This suggests that understanding the mechanisms underlying metabolic adaptations will allow us to develop precision medicine for the patients suffering from atherosclerosis.
MACROPHAGE SUBSETS
Generally, monocyte-driven macrophages are the most abundant immune cells, which can be trained by epigenetic reprogramming and metabolic rewiring to elicit a pro-inflammatory response in the atherosclerotic lesion.16 Distinct macrophage phenotypes regulate both plaque progression and stability of atherosclerotic lesions.17 Macrophage phenotype switching occurs in the region within the plaque in order to respond to the local microenvironment. It is known that M1 macrophages are predominately associated with an apoptotic phenotype and are counterbalanced by M2 macrophages. Hence, plaque instability may be the result of an imbalance in the M1 to M2 ratio. A novel macrophage phenotype called macrophage induced by oxidized phospholipids (Mox) is induced by oxidized phospholipids similar to what occurs during the atherosclerotic condition. This condition is characterized by an abundance of Nrf2-mediated redox-regulatory genes with reduced phagocytic and chemotactic capacities. Mox macrophages comprise 30% of macrophages in advanced atherosclerosis models.18 Based on scRNA-seq, mass cytometry, and fluorescence activated cell sorting, known as multi-plex immune phenotyping, 11–13 principal leukocyte clusters show high heterogeneities depending on the spatial lesions of the atherosclerotic plaques; in contrast, fewer diverse subsets are found in healthy aortas.19 In addition, 3 distinct macrophages were identified: resident macrophages, inflammatory macrophages, and a novel trigger receptor expressed on myeloid cells 2 (Table 1).15 A recent approach using atherosclerotic bone marrow (BM) chimeras in Ldlr−/− mice labeled with C-X-C chemokine receptor (CXCR) 1+: TdTomato cells allowed the authors to identify novel stem-like signatures in both progression and regression areas, which was simultaneously confirmed by other transcriptome analysis. These data demonstrate that macrophages under differing conditions reflect the plasticity integrated into the complex heterogeneity seen in the development of the atherosclerosis.2021 Also, Kim et al.21 reported that the scRNA-seq profile in nonfoamy plaque macrophages is associated with proinflammatory transcripts compared to foamy macrophages of the atherosclerotic intima. Hypercholesterolemia promotes monocytosis in BM derived hematopoietic stem and progenitor cells affected by epigenetic changes characteristic of long-lived, cell intrinsic memory.22 Furthermore, macrophages from atherosclerosis that are induced by a high fat diet exhibit higher cholesterol and cholesterol ester accumulation, which is positively associated with LDL levels. This results in the accumulation of glycerophosphocholine, unlike the increased phosphocholine in lipid loaded monocytes due to differential gene expression patterns.11 Excess cholesterol has multiple effects on clonal hematopoiesis and lesion inflammation in atherosclerosis.
Table 1
Characteristics of distinct macrophages in atherosclerotic lesions

Mox, macrophage induced by oxidized phospholipids; H3K4me3, histone 3 lysine 4 trimethylation; H3K27me3, histone H3 lysine 27 trimethylation; HDAC, histone deacetylase; TET2, tet methylcytosine dioxygenase 2; NLRP3, nucleotide-binding domain-like receptor protein 3; IL, interleukin; FAO, fatty acid oxidation; AMPK, AMP-activated protein kinase; PDK, pyruvate dehydrogenase kinase.
![]()
Epigenetic remodeling that is associated with lower histone H3 lysine 4 trimethylation (H3K4me3) and H3K27 trimethylation on the tumor necrosis factor (TNF)-α promoter leads to pro-inflammatory phenotypes in severe symptomatic coronary atherosclerosis patients versus control subjects.23 The other histone deacetylase 3 inhibitors in Ldlr−/− mice fed a Western diet ameliorated the atherosclerotic lesions by increasing the expression of ABCA1 and ABCG1.24 Likewise, oxLDL-induced long-lasting proinflammatory phenotypes are mediated by H3K4me3 position on proinflammatory cytokine promoters during foam cell formation.23 A deficiency of the epigenetic modifier enzyme tet methylcytosine dioxygenase 2 (TET2) correlates with increased somatic mutation frequency in hematopoietic cells. This leads to a remarkable increase in atherosclerotic plaque size, which in turn results in increased NLRP3 inflammasome-mediated IL-1β.25 Additionally, Fuster et al.25 showed that unregulated P-selectin in a TET2-deficient BMT model was abrogated upon MCC950 treatment. Unregulated P-selection acted as a NLRP3 inflammasome inhibitor and contributed to monocyte recruitment at the atherosclerotic plaques, followed by endothelial cell activation, which suggests that TET2 suppresses inflammasome priming. MCC950, a potent and selective NLRP3 inflammasome inhibitor, administrated in apoE−/− mice significantly attenuated atherosclerotic lesion development, consistent with a reduction in the number of macrophages inside the lesion. There was no change observed in the amount of collagen produced, or in the size of the necrotic core. Although IL-1β mRNA expression was not affected, MCC950 completely suppressed the secretion of IL-1β in both BM-derived macrophage and BM-derived DCs.26 IL-1β blockade caused by inhibition of the NLRP3 inflammasome may be a therapeutic target for the prevention and treatment of cardiovascular diseases in human subjects carrying clonal hematopoiesis-mediated disturbances.
Macrophage polarization requires metabolic reprogramming, which includes enhanced glycolysis and repurposing of mitochondrial function.272829 The enhanced markers of alternative activation are significantly associated with increased mitochondrial respiration, supported by higher levels of fatty acid oxidation (FAO) and AMP-activated protein kinase (AMPK) and lower levels of glycolytic enzymes in miR33-treated Western diet fed Ldlr/− mice. This resulted in the reduction of the atherosclerotic plaque size.30 Hypercholesterolemia reduced mitochondrial mass and maximized mitochondrial respiration, which may be responsible for the apoptosis-mediated necrotic core development by “foam cell” macrophages and is not attributed to typical inflammatory responses.31 The nuclear receptor Nr4a1 (Nur77) is required for the conversion of Ly6Chigh monocytes to Ly6Clow monocytes and is characterized by resolving macrophages for the tissue repair phase.32 Epigenetic changes in monocytes exposed to LDL affect the primed cells followed by upregulation of inflammatory cytokines associated with H3K4me3 in gene promoters.33 A high cholesterol/fat diet is associated with down-regulation of the cholesterol synthesis pathway. However, this pathway also enhances the innate immune responses to lipopolysaccharide by functionally reprogramming myeloid precursor cells.34 Our previous work has focused on the importance of the regulation of the pyruvate dehydrogenase complex by pyruvate dehydrogenase kinases (PDKs) in low grade inflammation-mediated obesity, insulin resistance, diabetes, and neurological disorders.35 These studies have shown that PDKs are a viable target for metabolic dysfunction under these conditions. Taken together, recent emerging data have demonstrated that metabolic reprogramming, from oxidative phosphorylation to aerobic glycolysis and/or FAO, determine the phenotype of macrophages in different conditions, which is still not characterized in atherosclerosis development.
ADAPTIVE AND CELLULAR IMMUNITY OF THE T CELL
The adaptive immune system can either promote or suppress inflammatory responses by distinct T cell subtypes.36 LDL-reactive CD4+ Th cell subsets have been identified in atherosclerosis by antigen-presenting cells (APCs). These cells are regulated by an LDL epitope via heavy oxidation and are categorized in the following groups: 1) pro-atherosclerotic Th1 cells regulated by T-box transcription factor 21/T-bet that secrete IFN-γ, which make the atherosclerotic plaque more vulnerable to rupture; 2) pro- and anti-atherosclerotic Th2 cells that secrete IL-4 and IL-5/IL-13/IL-33, respectively; 3) Th17 cells, as well as mast cells, and neutrophils that secrete IL-17A, which have conflicting effects on atherosclerosis but promote plaque stability and prevent cardiovascular events; and 4) protective regulatory T cells (Tregs), which are regulated by both forkhead box P3 (FOXP3) and transforming growth factor beta and act on the plaque by stabilizing the stimulation of collagen production (summarized in Table 2).710 An increase in the ratio of effector T cells (Teff) to Treg cells, controlled by T-bet and FOXP3, respectively, is a consequence of environmental cues in addition to co-existing Th1-Treg cells derived from previous Treg cells, leading to atherosclerotic lesion development.6 Likewise, Teff cells undergo changeover and can differentiate into Th17 cells by secreting IL-17A and IL-10 into pathological cytokines like IL-17A, IL-22, and IFN-γ. These are promoted by IL-1, IL-6, and IL-23, which are expressed in atherosclerotic lesions.3738 Recently, Zeboudj et al.39 showed that the epidermal growth factor receptor expressed in CD4+ T cells might be a target for atherosclerosis, a T cell-mediated vascular chronic inflammatory disease due to interference with T cell proliferation. The intrinsic roles of activated CD4+ T cells in both atherosclerosis and angiogenesis were investigated by identifying factors that stimulate collateral artery growth without aggravating atherosclerosis. This was done using a hind limb ischemic model to restore blood flow to the tissue, which is mediated by a lower accumulation of T cells.40 Systemic autoimmune disorders including psoriasis, systemic lupus erythematosus (SLE), and rheumatoid arthritis are thought to be associated with autoreactive T cells, including Th17 cells, and/or follicular helper T cells (TFH).41 Furthermore, a pro-atherogenic environment promoted B-cell stimulatory functions on TFH cells in lupus-prone mice and the accumulation of CD4−CD11b+ DCs, with an increased capacity for IL-27 production.41 Impaired cholesterol efflux is significantly associated with SLE. SLE is characterized by enlargement of the lymph node and glomerulonephritis and Th1 and Th17 polarization in DC cholesterol accumulation, which is predominantly mediated by increased IL-1β secretion and upregulated granulocyte macrophage colony-stimulating factor beta expression.42
Table 2
Characteristics of diverse T cells in atherosclerotic development

Th, T helper type; Treg, regulatory T cell; FAO, fatty acid oxidation; mTOR, mammalian target of rapamycin; FOXP3, forkhead box P3; TGF, transforming growth factor; IL, interleukin; IFN, interferon; AMPK, AMP-activated protein kinase.
![]()
Resident T cells in the liver, which is the major organ where dietary cholesterol is processed and lipoproteins are synthesized, are relocated into the atherosclerotic lesions with different ratios of Teff and Treg cells.6 Interestingly, Treg-specific ABCG1 deficiency in an atherosclerosis model exhibits more Treg cells and fewer atherosclerotic lesions compared to the control. It is supported by the observation that more cholesterol in the lysosome interferes with mammalian target of rapamycin (mTOR)-mediated signal transducer and activator of transcription (STAT) 5 signaling in ABCG1 deficient T cells.43 Distinct T cell subsets depend on many different metabolic features: 1) resting naïve T cells utilize the tricarboxylic acid cycle in order to generate ATP from glucose-derived pyruvate, lipids, and amino acids; 2) antigen-induced clonal proliferating T cells exhibit aerobic glycolysis, lipid synthesis, and reduced lipid oxidation; 3) resting T memory cells exclusively re-activate the TCA cycle using lipid sources; 4) effector Th1/Th17 and Th2 require aerobic glycolysis and are mediated by mTOR1 and mTOR2 activities, respectively; and 5) Treg is regulated by AMPK and FAO.44 The different fates of T cell subsets are significantly dictated by mitochondrial dynamics due to fusion and fission events in order to meet metabolic needs during T cell activation.45 C-C chemokine receptor type 5 Teff cells are identified as atherosclerosis-specific CD4+ T cells in the aorta and para-aortic lymph nodes with a unique combination of surface markers, CD4+CD25−CD44highDC62Llow with FOXP3+T-bet+. These cells secrete IFN-γ, IL-2, IL-10, and TNF, similar to Teff cells.46 Intracellular cholesterol accumulation in T cells promotes Treg cell differentiation through ABCG1-specific deficiency, which leads to reduced T cell activation and attenuated atherosclerotic lesions.43 Induced hypoxia-inducible factor-1α in APCs interferes with T cell proliferation and Th1 polarization, mediated by STAT3-IL12, and attenuates atherosclerosis.47 Peripheral tolerance in a low glucose and high lactate environment is mediated by the expression of FOXP3 in Treg cells, which downregulates Myc and glycolysis and upregulates the conversion of pyruvate from lactate, followed by oxidative phosphorylation, and vice versa.48 The cholinergic anti-inflammatory pathway involves the release of acetylcholine from T cells, which binds to nicotinic acetylcholine receptors on immune cells in the spleen. This process may alleviate atherosclerosis as well as hypertension by reducing proinflammatory cytokines such as TNF-α and IL-17.49 In human immune system-reconstituted mice, which differs from the adaptive immune system in inbred mouse strains, moderately increased cholesterol can disrupt human T cell homeostasis in vivo, which contributes to T cell-mediated inflammatory diseases.2
HUMORAL IMMUNITY OF B-CELL AND AUTOIMMUNE DISEASE
Based on both splenectomy and B cell transfer experiments in atherosclerotic mice and splenectomized patients, it is clear that B cells play a role in atherosclerosis, but more studies are needed to determine the mechanisms involved.7 A drug-based immunotherapy that can interfere with the B cell activating factor (BAFF)/BAFF receptor axis due to antibody binding to oxLDL, phospholipid, and β2GPI/oxLDL, prevents atherosclerotic vasculopathy under autoimmune conditions.50 The TFH-germinal center B-cell axis accelerates atherosclerosis, which is suppressed by CD8+ Tregs in tertiary lymphoid organ development.51 Increased pathogenic subclasses of immunoglobulin G by B cells in atherogenic conditions can be explained by hyperlipidemia-toll-like receptor 4-IL27-CXCR3 in the TFH cell axis.41 Although the number of patients with autoimmune diseases, including rheumatoid arthritis and psoriasis, carry increased risk for cardiovascular diseases, cytokine-targeted therapies involving TNF-α or IL-17 antibodies remain unexplored for reducing plaque inflammation and progression.6 Likewise, dyslipidemia in chronic inflammatory diseases may be associated with myocardial infarction risk in patients diagnosed with rheumatoid arthritis.52 Leukocytosis in myeloproliferative disease is positively associated with the incidence of atherosclerosis, contributed by hypercholesterolemia dependent- and Glut1-mediated glucose uptake-monocytosis.653 The underlying mechanisms of humoral immunity and autoimmune diseases promoting atherosclerosis remain unexplored.
CONCLUSION
Chronic low-grade inflammation in multiple organs increases the risk of developing obesity, diabetes, cardiovascular diseases, and cancers, indicating a major role for the immune system in the etiology of metabolic disorders associated with atherosclerosis risk. Recruitment of M1 macrophages, IFN-γ-secreting Th1 cells, CD8+ T cells, and B cells in atherosclerotic lesions drives the inflammatory response, locally promoting systemic inflammation. The activation of immune cells mediated by metabolic reprogramming has been intensively investigated for developing therapeutic targets for cardiovascular diseases, including atherosclerosis.
 XML Download
XML Download