

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Various emotional states exist as the human nervous system has been developed so as to react appropriately to external stimuli. Russell's model,1 which is widely used as a reference for emotional distinction, classifies emotional states according to two dimensions, arousal and valence. The influence of valence and arousal on cortical information processing has been extensively studied. Emotion induced by visual stimuli provides an opportunity to explore emotional information processing. A number of previous studies have investigated the cortical information processing process in response to visual emotional stimuli.2345678 Furthermore, it has been pursued to understand the changes of emotional information processing according to neuropsychiatric disorders such as major depressive disorder and alexithymia,9 as well as degenerative brain disease.10
Emotional visual stimuli induce not only emotional information processing but also visuoperceptual processing.11 Valence is a central feature of emotion, contrasting the effects of positive versus negative states. Arousal is the physiological state of being awoken. The dimension of arousal ranges from calming to exciting. It is not completely revealed how the valence and arousal affect these two procedures. Emotional information draws attention immediately (within less than around 200 ms), and initial sensory information processing is usually enhanced by emotional stimuli compared to neutral ones.12 Both arousal and valence of the stimuli may affect sensory processing, as it has been demonstrated that the stimuli with high arousal and low valence induce significantly enhanced brain activities.712 On the other hand, emotional stimuli are motivationally relevant, and thus, cause strong memory encoding even when the encoding is not intended.13 It is well known that both arousal and valence are involved in herein.31415
Spatiotemporal functional neuroimaging technology is required to quantitatively characterize the effects of valence and arousal dimension on the constituent steps of visual emotional information processing. Noninvasive functional brain mapping studies have been performed on this issue using either electroencephalogram (EEG), magnetoencephalogram (MEG), or functional magnetic resonance imaging (fMRI).16 However, dynamic spatiotemporal neuroimaging is necessary in order to overcome the inherent limitations in spatial (EEG/MEG) and temporal (fMRI) resolution and to reveal neural substrates and their temporal behaviors.
The purpose of this study is to quantitatively investigate the effects of valence and arousal on spatiotemporal cortical information processing of emotional visual stimuli. Overall cognitive process can be divided into several units which may be based on overlapping spatial and temporal neural substrates. Distributed source localization analysis of EEG was adopted to characterize the spatiotemporal pattern of event-related cortical current density (ERCD) in response to emotional visual stimulation. A nonparametric cluster-based permutation test17 was used to find meaningful time and space without a priori knowledge. This enables us to find the spatiotemporal clusters showing significant differences between emotional states, based on the assumption that meaningful information processing units have spatially and temporally localized neural substrates.
Here our focus was on visuoperceptual analysis and memory encoding among the many constituent units of overall cortical processing. The memory encoding is expected to be specifically affected by arousal level, since stimuli evoking higher arousal are motivationally relevant, and cortical regions within the parietal memory network,18 which are found to be involved in memory encoding and retrieval processes, is expected to be involved here. The stimuli with positive/negative valence are expected to result in enhanced visuoperceptual processing compared to neutral ones. In particular, negative valence would cause enhanced visuoperceptual processing, presumably within inferior temporal cortical activity, presumably due to visual morphological feature processing during the memory encoding which is unintentionally, but automatically induced.1920 The results of this study may provide valuable information for understanding the changes of cortical function according to neuropsychiatric disorders.
METHODS
Participants
Nineteen healthy men without any history of neurological and psychiatric illnesses participated in the study (23.18 ± 1.92 years). Only men subjects were recruited to minimize the impact of potentially confounding variable according to gender differences in the cortical processing of visual emotional stimuli, (which is not direct interest of our study).21 All subjects were undergraduate students of Yonsei University and had normal or corrected-to-normal vision.
Visual stimuli
The visual stimuli were constructed from the International Affective Picture System (IAPS).22 The selection of images was performed based on the valence and arousal levels reported by the participants. We selected 100 images that may induce positive emotion (valence, 6.92 ± 0.56; arousal, 5.52 ± 0.75), 100 images that may induce negative emotion (valence, 2.48 ± 0.45; arousal, 5.66 ± 0.72), and 200 non-emotional pictures (valence, 4.97 ± 0.51; arousal, 3.65 ± 1.04). The IAPS stimuli used in this experiment for each condition are specified in Table 1. The number of neutral stimuli was set to be equal to the sum of positive and negative stimuli, so that the numbers of emotional and neutral stimuli were matched. Valence levels of the two emotional categories (positive and negative) were found to be significantly different (independent t-test, t(198) = 69.78, P < 0.001, Bonferroni corrected), but there was no significant difference in arousal levels of them (independent t-test, t(198) = −1.60, P = 0.330, Bonferroni corrected).
Table 1
The number of IAPS pictures used in the experiment

Experimental procedure
The experiment task was divided into four blocks of 12.11 ± 0.25 minutes. Each block consisted of 100 trials, including 25 positive, 25 negative, and 50 neutral visual stimuli. Five-minute resting periods were given between two blocks. Neutral pictures were always presented between emotional pictures, in order to avoid the effect of emotion induced by preceding stimulus. Fig. 1 shows the experimental task and the temporal structure of a single trial. The pictures were presented for 2,000 ms, which was set as the epoch of interest for the analysis. A fixation mark (‘+’) was presented for 1,000 ms before each stimulus. The subjects were instructed to determine whether they liked or disliked the visual stimulus as soon as possible, but to keep the intention of preference (like or dislike) for another 2,000 ms after the stimuli was extinguished while the fixation mark was displayed. Then, the subjects were to respond whether they ‘liked’ or ‘disliked’ by either ‘○’ or ‘×’ as shown in Fig. 1 as soon as possible within 2,000 ms. The location of ‘○’ and ‘×’ was randomized in order to minimize the prediction of the direction, and motor preparation.
EEG recording and preprocessing
EEGs were recorded at a sampling rate of 500 Hz using a 64 channel Ag/AgCl electrode cap (actiCAP; Brain Products, GmbH, Munich, Germany) according to the extended 10-20 system. The impedances of all electrodes were kept below 10k ohm. The reference and ground electrodes were placed at FCz and AFz, respectively. A bandpass filter (0.03–100 Hz) and 60 Hz notch filter was applied. EEGLAB was used for the preprocessing of brain potential.23 The single-trial waveforms were excluded from further analysis if they were severely contaminated by drift or transient artifacts due to head/electrode motion. Since we had the knowledge on the ground truth of valence of each visual stimulus, we further excluded the trials in which the subject's response was apparently incorrect. Each image in the International Affective Picture System (IAPS) has official normative rating on emotional condition. The rating can be considered to reflect normal emotional response (i.e., ground truth), and thus, each image can be classified into positive/negative/neutral states based on the rating. The subjects were asked to report whether they liked or disliked the stimuli. A trial was excluded from further analysis, if the response was not in accordance with the emotional state which was determined from the rating. For example, when a subject answered that he liked an apparently negative picture, we considered he showed abnormal emotional response due to some reason including abnormal personal preference, personal experience, or inattentiveness to the stimulus. The number of remaining trials was 89.05 ± 13.88, 88.37 ± 7.50, 187.63 ± 14.56 for positive, negative and neutral stimuli, respectively. Stereotyped artifacts such as eye movement or EMG were removed using independent component analysis (ICA).24 Subsequently, the EEGs were re-referenced according to the average reference.
Source localization
An open source MATLAB toolbox Brainstorm was used to calculate the cortical current source density on cortical surface.25 Boundary elements method was used for the forward problem. International Consortium for Brain Mapping (ICBM) 152 template was used as default anatomy.26 The head model was computed with a symmetric boundary element method (BEM) using OpenMEEG implemented in Brainstorm toolbox.27 The number of vertices in scalp, outer skull, inner skull layer were all set to 1922, and the conductivities of them were set to 1, 0.0125, 1 respectively. The thickness of skull was set to 4 mm. The current density at 15,002 points on cortical surface was calculated by solving the inverse problem with weighted minimum norm estimation.28 The cortical surface was homogeneously parcellated into 500 points using surface clustering implemented in Brainstorm toolbox,29 for the whole-brain analysis without prior knowledge. Finally, current density time-series on 500 cortical points were obtained.
Statistical analysis
Conventional analysis of event-related potential (ERP) utilizes prior knowledge of electrodes and time which are highly likely to show significant difference between emotional conditions. However, our purpose here is to find out the critical time and cortical region from the ERCD time-series itself, without using prior knowledge. This requires statistical comparisons of cortical current density at many points in time-cortex space, resulting in a multiple comparison problem involving a very large number of simultaneous comparisons. In our case here, there exist 250,000 (= 500 vertices × 500 time points) multiple comparisons. We adopted a nonparametric cluster-based permutation test17 to deal with this problem. As shown in Fig. 2A and B, this method tries to find a set of continuous spatiotemporal points which show significant changes in ERCD according to conditions, which is referred to as a ‘cluster.’17
Fig. 2
Illustration of the procedure for finding spatiotemporal regions correspoding to significant differences between conditions using nonparametric cluster-based permutation test. (A) Calculation of univariate test-statistics at all the points in time-vertex space. (B) Left: selected regions in time-vertex space with high test-statistics. If two points are continuous with respect to either time and vertex, they are grouped as a cluster as shown. Three clusters with P < 0.05 are shown, along with the sum of test-statistics within each cluster. Right: The clusters with high sum of test-statistics are determined based on the null distribution obtained from random permutation.
ERCD = event-related current density.

The detailed procedure is as follows: 1) ERCD averaged over all trials at all 500 cortical points, during −100 to 1,000 ms (550 time samples) are calculated; 2) Baseline correction of the averaged ERCD time-series is performed by subtracting average ERCD within −100 to 0 ms; 3) Statistical comparisons of three valence conditions are performed by one-way ANOVAs at 500 × 500 points in time-cortex space (500 temporal points, excluding 50 points within −100 to 0 ms baseline period); 4) The points with large F values, corresponding to P < 0.05, are selected; 5) The clusters of critical spatiotemporal points are defined by combining the points if two significant points are consecutive in time or within 30 mm in space17; 6) The ‘mass’ of a cluster is defined as the sum of F values within the cluster; 7) Surrogate data are obtained by random permutation of three conditions within each subject (5,000 times), and the null distribution of cluster mass is obtained by repeating steps 1)–6) above for the surrogate data; 8) The cluster masses found in step 6) are compared to the highest 5% of null distribution in step; 7) to determine whether they are significantly large, i.e., whether corresponding point corresponds shows significant difference between emotional conditions.
For the nonparametric cluster-based permutation test described above, the Mass Univariate Toolbox of Groppe et al.30 was modified to enable the ANOVA in addition to the t-test. The temporal epoch for the analysis was set to 400–800 ms, according to the earliest and latest time in the significant clusters determined as described above. The significant clusters were further parcellated to anatomical regions following the automated anatomical labeling (AAL) of Tzourio-Mazoyer et al.31 after taking time-average over the temporal epoch described above. The ERCDs of the vertices belonging to each parcel, i.e., each region of interest (ROI), were averaged to obtain ERCD time series for the parcel. Here the clusters were grouped to 6 ROIs (among 116 in the AAL database31) including bilateral central, cingulate, and inferior temporal areas.
Finally, two-way repeated-measures ANOVA was performed to analyze averaged ERCD amplitude over 400–800 ms of each region of interest. Within-subject factors included emotional conditions (3 levels) and ROIs (6 levels). Post-hoc t-test was performed to compare ERCDs between emotional conditions for each ROI. The P value was corrected by false discovery rate (FDR)32 method, proposed by Benjamini and Yekutieli.33 Statistical results of ROIs which showed significant ERCD differences among the conditions were displayed for simplicity.
RESULTS
Fig. 3A and B shows a scatter plot of the valence and arousal levels of all visual stimuli and statistical comparison of the ratings of valence and arousal levels among positive, negative, and neutral categories. Valence level was significantly different among the three categories (F [2,397] = 2,438.63; P < 0.001), and all the post-hoc pairwise comparisons between two categories were significantly (positive-negative, P < 0.001; positive-neutral, P < 0.001; negative-neutral, P < 0.001; Bonferroni corrected) as shown in Fig. 3B. Arousal levels were also significantly different among categories (F [2,397] = 263.58, P < 0.001). Post-hoc t-test showed that the arousal levels were not significantly different between the positive and negative categories (P = 0.33, Bonferroni corrected), whereas they were significantly higher for the positive/negative as compared to the neutral stimuli (positive-neutral, P < 0.001; negative-neutral, P < 0.001; Bonferroni corrected).
Fig. 3
Arousal and valence levels of emotional stimuli. (A) Distribution of arousal and valence levels of visual stimuli. (B) Statistical comparison of the valence and arousal level (***P < 0.001, Bonferroni corrected, independent t-test).

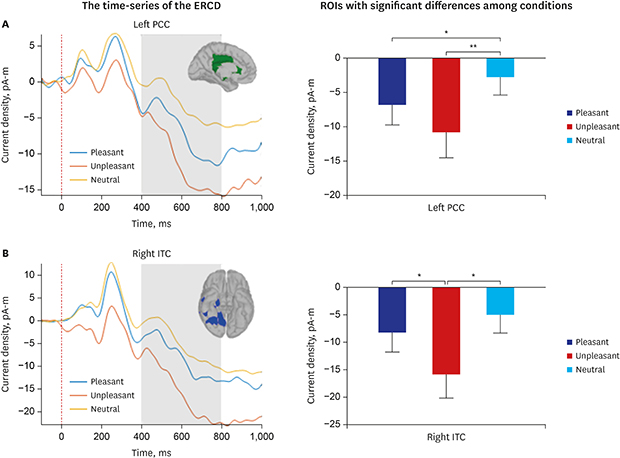
Fig. 4 shows the cortical regions showing significant ERCD differences among the three emotion conditions in 400-800 ms. Among the six ROIs determined by the cluster-based permutation test, only two ROIs showing statistical difference between categories are presented in Fig. 4 for simplicity (left PCC [green] and right IT [blue]). Significant main effects of ROI (F [5,90] = 2.719; P = 0.025) and emotional category (F [2,36] = 7.977; P = 0.010) were found by repeated-measures ANOVA, and the interaction between the two factors was also statistically significant (F [10,180] = 2.261; P = 0.016).
Fig. 4
Cortical regions showing significant event-related current density differences between the three emotion conditions. Among the six region of interests which were determined by the cluster-based permutation test, only two are displayed for simplicity (left PCC [green] and right ITC [blue]).
PCC = posterior cingulate cortex, ITC = inferior temporal cortex.

Significant changes of ERCD between emotional categories were found only in the two ROIs presented in Fig. 4 (left PCC and right IT). In the left PCC, both positive and negative stimuli showed more negative ERCD compared to the neutral ones (Fig. 5A). The ERCD for the positive was significantly more negative than that for the neutral (t [18] = −2.936; P = 0.013; FDR corrected), and that for the negative was also significantly more negative than that for the neutral (t [18] = −3.870; P = 0.003; FDR corrected). However, it was not significantly different between the positive and negative categories (t [18] = 1.836; P = 0.083; FDR corrected).
Fig. 5
Spatiotemporal characteristics of the two ROIs (left PCC and right ITC). (A) The time-series of the ERCD at the left PCC and its statistical comaprison among emotional categories. (B) The time-series of the ERCD at the right ITC and its statistical comaprison among emotional categories. Error bars denote standard errors.
ERCD = event-related current density, PCC = posterior cingulate cortex, ROI = region of interest, ITC = inferior temporal cortex.
*P < 0.05; **P < 0.01; pairwise t-test, FDR corrected.

In the right IT, it was remarkable that the ERCD was significantly more negative for the negative compared to the other two categories (Negative-positive: t [18] = −2.545; P = 0.030; FDR corrected. Negative-neutral: t [18] = −3.096; P = 0.018; FDR corrected) (Fig. 5B), whereas it was not significantly different between the positive and neutral categories (t [18] = −1.488; P = 1.000; FDR corrected).
DISCUSSION
In this study, cortical information processing of emotional visual stimuli was investigated using distributed source analysis of multichannel event-related EEG. Overall information processing includes visual pattern analysis as well as emotional analysis, and we expected that the valence and arousal would have different effects on each of these two steps. The most noticeable differences between emotional categories were found at 400−800 ms interval in posterior cingulate and inferior temporal cortices. The former and the latter were predominantly affected by arousal and valence, respectively. We found that arousal has strong effect on episodic memory encoding. As for the valence, only the negative one seems to have a large influence on cortical activities in visual cortical area. The anatomical locations of the neural substrates of emotional visual stimulus processing were determined by EEG source imaging, which has inherent limitation in spatial resolution.28 However, our results are in accordance with current knowledge on functional neuroanatomy.343536
According to previous ERP studies on emotional visual stimuli processing, the interpretation of the meaning of stimuli occurs after 400 ms, especially for the encoding of episodic memory on motivationally relevant stimuli.13 Novel stimuli are quickly and automatically memorized without efforts, which is important characteristic of episodic memory. Cognitive process of episodic memory embraces encoding, consolidation, storage, and retrieval. For the encoding, new information is combined with previous memories, and thus, it is expected that recollective interpretation of incoming visual stimuli at 400−800 ms is closely related to the memory encoding.
The difference in cortical activities according to arousal level was observed predominantly at 400−800 ms in PCC. Considering the functional role of PCC, the result suggests that arousal influences automatic, unintended memory encoding of arousing visual stimuli. Previous studies have shown that the PCC is activated during episodic memory retrieval,34 and, is involved in both memory encoding and retrieval, along with precuneus, posterior inferior parietal lobule. These regions together are suggested as a parietal memory network.18 An fMRI study on visual presentation of emotional words showed that left PCC is selectively activated by arousing stimuli, and not affected by valence level,35 in accordance with our results.
The activities in right inferior temporal cortex at 400−800 ms epoch in response to the stimuli with negative valence are expected to be due to the evaluation of the risk associated with the negative-valence stimuli.37 It is interesting that the activities of the visual area at this later period, which seems to be after the end of earlier visual pattern analysis, are increased for the negative stimuli. The IT cortex belongs to the ventral stream of visual information processing which is involved within recognizing patterns, faces, and objects.36 The sensitivity of the IT on negative stimuli may also be due to its connectivity with amygdala,38 which is a part of cortical circuits for the negative stimuli processing.39
In conclusion, we found that arousal has strong effect on episodic memory encoding, and that only negative valence seems to have large influence on cortical activities for the visual analysis, at relatively late epoch (400−800 ms). These results may provide valuable information for understanding the changes of cortical function according to neuropsychiatric disorders.
 XML Download
XML Download