

 ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The primary role of sebocytes is the making sebum, the specialized lipid complex, that covers the surface of skin thereby providing a barrier function1. Sebum production is controlled by various factors such as androgens, corticotropin-releasing hormone, vitamin D and insulin-like growth factor-1 (IGF-1)2. At puberty, sebum production increases in accordance with the increase of circulating androgens, and the resultant sebum increase often leads to the development of acne. When sebocytes produce excess sebum, the microenvironment surrounding hair follicle changes into lipid-rich hypoxic condition that enables the flourishing of lipidophilic commensal bacteria such as Propionibacterium acnes. In addition, the metabolites produced by microbes trigger the activation of perifollicular inflammation, exacerbating the acne condition34.
Currently, many drugs are employed for treatment of acne. For example, the antibiotics such as tetracycline, doxycycline and minocycline are used for controlling microbes. In addition, vitamin A derivatives such as adapalene, isotretinoin and tazarotene are used for inhibiting the hyperkeratinization of pilosebaceous duct5. Although many drugs are widely used for treatment of acne, there is still a demand for the development of new treatments that have little adverse effects and high efficacy. Since the excess production of sebum is the starting event in pathogenesis of acne, targeting the sebum production of sebocytes is a potential strategy for acne therapy.
In a previous screening test using cultured human sebocytes, we found that bilobetin had strong anti-lipogenic potential. In this study, we demonstrate that bilobetin inhibits IGF-1–induced sebum production, suggesting that bilobetin can be developed for acne treatment.
MATERIALS AND METHODS
Ethics statement
Skin tissues were obtained from volunteers under the written informed consent, and the present study was approved from the ethical committee of the Institutional Review Board of Chungnam National University Hospital (IRB no. 1011-135, 2016-07-009).
Cell culture
Human scalp tissues were immersed briefly in 70% ethanol for sterilization, then sebaceous glands were separated using microforcep under the stereomicroscope. Isolated sebaceous glands were attached to the culture dish, and incubated in Sebomed® medium (Biochrom, Berlin, Germany) supplemented with 10% fetal bovine serum and 5 ng/ml recombinant human epidermal growth factor (Life Technologies Corporation, Grand Island, NY, USA). Usually, sebocytes were grown out from explanted sebaceous glands in 1 week.
For a long and continuous maintenance of sebocytes cell line, we immortalized the primary cultured cells using a recombinant retrovirus expressing simian virus 40 T (SV40T) antigen. The retroviral vector pLXIN-SV40T was transfected into the retrovirus packaging cell line PT67 (Clontech Laboratories, Mountain View, CA, USA). The medium containing retrovirus was harvested, filtrated through a 0.22-µm low protein binding filter (Merck KGaA, Darmstadt, Germany), and then transferred to primary cultured sebocytes. After incubation overnight, the retrovirus-containing medium was removed and fresh medium was added. The immortalized sebocytes were selected in medium containing 200 µg/ml of G418 (Sigma, St. Louis, MO, USA) for 4 weeks.
Cell viability test
SV40T-transformed sebocytes (SV-sebocytes) were seeded in 6-well plate at a density of 2×105 cells/well, treated with bilobetin (Biopurify Phytochemicals Ltd., Chengdu, China) for 1 day. Cell viability was measured by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay.
Cell growth assay
To measure the cell growth, [3H]thymidine uptake assay was carried out. SV-sebocytes were seeded in 6-well plate and treated with bilobetin for 2 days in the presence of 1 µCi [3H]thymidine (Amersham, UK). Cells were lysed with 0.1 N NaOH, then radioactivity was measured using a liquid scintillation counter.
Oil Red O staining
SV-sebocytes were grown on cover glass and treated with bilobetin. After fixation with 10% formalin for 5 minutes, cells were stained with Oil Red O solution. Stained cells were vigorously washed with distilled water, and then counterstained with hematoxylin.
Lipogenesis assay
To check the lipogenesis, a well-established 14C-acetate incorporation assay was used6. SV-sebocytes were treated with bilobetin for 1 day, then 2 µCi of [1-14C]acetic acid (PerkinElmer, Boston, MA, USA) was added and incubated for a further 6 hours. Cellular lipids were extracted with the solvent comprising of chloroform and methanol (2:1). After evaporation of solvent, cellular lipids were dissolved in chloroform and separated using a thin layer chromatography (TLC silica gel 60 F254; Merck KGaA). The developing buffer for TLC consists of hexane and ethyl acetate (6:1). Lipids were visualized by autoradiography.
Luciferase reporter assay
The squalene synthase-luc reporter adenovirus (Ad/SS-luc) was produced previously7. For luciferase reporter assay, SV-sebocytes were seeded in 12-well plate and infected with Ad/SS-luc for 6 hours. After removal of adenovirus-containing medium, cells received fresh medium containing IGF-1 and bilobetin for 1 day. Luciferase activity was measured using the dual luciferase reporter assay system (Promega, Madison, WI, USA).
Western blot
Cellular extracts were prepared using a lysis buffer (Intron, Daejeon, Korea). After sodium dodecyl sulfate–polyacrylamide gel electrophoresis, proteins were transferred to nitrocellulose membranes. The membranes were incubated with primary antibody, and then sequentially incubated with peroxidase-conjugated secondary antibodies. The Western bands were obtained by enhanced chemiluminescence (Intron), and quantified using ImageJ software (ver. 1.50i). The following primary antibodies were used: sterol response element binding protein-1 (SREBP-1), actin (Santa Cruz, CA, USA); SREBP-2, squalene synthase (farnesyl-diphosphate farnesyltransferase 1, FDFT1) (Abcam, Cambridge, UK); stearoyl-CoA desaturase (SCD) (Thermo Scientific, Rockford, IL, USA); IGF-1-R, phospho-IGF-1-R, AKT, phospho-AKT, ERK1/2, phospho-ERK1/2 (Cell Signaling Technology, Beverly, MA, USA).
RESULTS
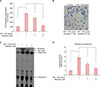
Bilobetin is a biflavonoid isolated from crude leaf extracts of ginkgo biloba8. It is made up of two structurally very similar but not identical flavonoid molecules by covalent bonding (Fig. 1A). To determine the cytotoxic effect, we treated SV-sebocytes with bilobetin at various concentrations. As a result, bilobetin did not show significant cytotoxicity up to the dose of 4 µM (Fig. 1B).
To investigate the effect of bilobetin on lipid production of sebocytes, we employed a well-established IGF-1–induced lipogenesis model69. Since IGF-1 stimulates sebocyte proliferation and lipogenesis10, we first examined the effect of bilobetin on cell growth using [3H]tymidine uptake assay. Treatment of SV-sebocytes with IGF-1 resulted in increase of cell growth, and pretreatment with bilobetin inhibited IGF-1–induced cell growth significantly (Fig. 2A). We next examined the effect of bilobetin on the production of neutral lipids by Oil Red O staining. IGF-1 treatment increased lipid production, and pretreatment with bilobetin resulted in significant inhibition of IGF-1–induced lipid production (Fig. 2B). To further analyze the lipid production of sebocytes, we performed TLC. Result showed that bilobetin markedly inhibited IGF-1–induced lipid production, including squalene and wax ester. In addition, bilobetin also blocked significantly the production of triglyceride and cholesterol (Fig. 2C). It has been well known that squalene and wax ester are the unique lipids found in sebum2, thus we examined whether bilobetin can affect the promoter activity of squalene synthase (FDFT1) gene. Cells were transduced with recombinant adenovirus harboring the squalene synthase promoter-luc cassette, then treated with IGF-1 and bilobetin. Consistent with data obtained from TLC, IGF-1 increased the squalene synthase promoter activity, and bilobetin significantly inhibited IGF-1–induced promoter activity (Fig. 2D). These results suggest that bilobetin may be applicable for specific inhibition of sebum production in sebocytes.
During the synthesis of lipids, various transcription factors and lipogenic enzymes are involved1112. We thus investigated whether bilobetin affected the expression of several lipogenic regulators. As expected, IGF-1 treatment led to increase of lipogenic transcription factors, SREBP-1 and SREBP-2, together with increase of lipogenic enzymes such as squalene synthase (FDFT1) and SCD. Pretreatment of bilobetin significantly inhibited IGF-1–induced up-regulation of lipogenic regulators (Fig. 3A). Since it has been demonstrated that IGF-1 induces SREBP-1 via the IGF-1 receptor (IGF-1-R) and downstream signaling pathway in sebocytes6, we examined the effect of bilobetin on intracellular signaling. When sebocytes were activated by IGF-1, the phosphorylation of IGF-1-R and AKT was significantly increased. Pretreatment with bilobetin did not affect the IGF-1–induced phosphorylation of IGF-1-R, while the phosphorylation of AKT was significantly inhibited by bilobetin. By contrast, phosphorylation of ERK1/2 induced by IGF-1 was not inhibited by bilobetin (Fig. 3B). These results suggest that action mechanism of bilobetin is primarily related to its inhibitory potential on AKT signaling.
DISCUSSION
Acne is a very prevalent skin disease, about 85% of young adults aged 12 to 25 years are affected by this skin problem13. Excessive sebum production and increased sebocyte proliferation play pivotal roles in the pathogenesis of acne14. In non-pathological condition, sebum constitutes the majority of skin surface lipids and exerts its role as a biological barrier by producing anti-microbial activity as well as a physico-chemical barrier of the skin15. However, excessive sebum makes the microenvironment surrounding hair follicle more favorable to colonization of lipophilic bacteria such as P. acne, thereby contributing to establishment of pathologic condition. In addition to the role for supporting bacterial growth, sebum may exert its role as a direct inducer for acne symptom. For example, squalene monohydroperoxide is the primary peroxidation product in human skin surface lipids that can induce hyperplasia and hyperkeratosis of the epithelium in follicular infundibulum16. Thus, targeting the sebum production in sebocytes is a good strategy for preventing acne. In this study, we demonstrated that bilobetin inhibited IGF-1–induced sebum production via the inhibition of AKT signaling in cultured human sebocytes.
Bilobetin is one of biflavonoids isolated from crude leaf extracts of ginkgo biloba8. Many biological activities of bilobetin have been reported. For example, bilobetin inhibited cAMP-phosphodiesterase in rat adipose tissue17. In other example, bilobetin ameliorates insulin resistance by protein kinase A-mediated phosphorylation of proliferator-activated receptor-α in rats fed a high-fat diet18. In this study, we demonstrated that bilobetin inhibited IGF-1–induced sebum production in sebocytes. Bilobetin significantly inhibited IGF-1–induced increase of lipogenic regulators including SREBP-1 and SREBP-2. Furthermore, bilobetin significantly blocked IGF-1–induced activation of AKT signaling. Since it has been demonstrated that IGF-1 signaling play an important role in sebum production through induction of lipogenic transcription factors such as SREBP-16, our results suggest that effects of bilobetin on sebum production may be due to its inhibitory potential on IGF-1–induced AKT signaling. As previously recognized, IGF-1 promotes seborrhea and inflammation, thereby playing an important role in the pathogenesis of acne24. Therefore, efficient blockage of this signaling is a good strategy in the treatment of acne, and bilobetin can be a candidate for this purpose. Nonetheless, IGF-1 is one factor affecting the sebum production, and there are still many important factors critically involved in pathogenesis of acne, such as androgen and commensals. Thus, the limitation of bilobetin should be considered in the context of developing the drugs for acne treatment.
In summary, we demonstrated that bilobetin inhibited IGF-1–induced sebum production in sebocytes through the inhibition of IGF-1–induced AKT signaling. Our results suggest that bilobetin can be applicable for acne treatment.


 XML Download
XML Download