

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Bone is a specialized form of connective tissue that is constantly being remodeled. It is formed via three sequential processes: bone extracellular matrix is synthesized by osteoblasts (osteogenesis); bone is resorpted by osteoclast to form its shape in response to the physical environment (modeling); and the milieu is changed to adjust to the demands of the growth (remodeling) [1]. Each of these processes is mediated by the balance of bone-forming osteoblasts and bone-resorbing osteoclasts [2]. A shift in this balance toward the activation of osteoclasts results in many clinical diseases, such as osteopenia and osteoporosis. Osteoporosis has been emerging as a global health concern that affects a wide range of populations, especially in old age [3]. Recently, osteoblast activity is considered a potential therapeutic target to control the osteopenic disease.
Osteoblasts are bone-forming cells that synthesize bone matrix, regulate mineralization, and eventually differentiate into osteocytes, or bone lining cells. They originate from mesenchymal stem cells and can differentiate into mature osteoblasts [4]. Mesenchymal stem cells give rise to preosteoblasts via activation of master transcriptional regulator, runt-related transcription factor 2 (Runx2) [5]. Thereafter, the growth and differentiation of osteoblast proceed in three stages with specific gene expression; the cells proliferate continuously and express collagen type I (COL1A1), TGF-β, fibronectin, and osteoponin (OPN); the extracellular matrix matures and expresses several extracellular matrix proteins including alkaline phosphatase (ALP) and COL1A1; and the matrix mineralizes and becomes enriched with osteocalcin, bone sialoprotein (BSP), and OPN. Subsequently, the mature osteoblasts with cuboidal shape is developed [67].
Remifentanil is a synthetic opioid acting on µ-opioid receptors, which is rapidly metabolized by nonspecific esterases in the plasma, giving it the characteristic of having very short onset and offset times [8]. Due to these properties, remifentanil has been used widely as an adjuvant analgesic drug during general anesthesia and sedation. The effect of remifentanil on osteoblasts has been investigated in previous studies. Yoon et al. [9] reported that remifentanil has a protective effect against oxidative stress on human osteoblasts. In another study, it was reported that pre-conditioning with remifentanil enhanced osteoblast maturation under hypoxia-reoxygenation conditions [10]. In the current study, we investigated the effect of remifentanil on osteoblast differentiation under normoxia state using C2C12 cells. We also investigated the mechanism of remifentanil-mediated osteoblast differentiation by analyzing osteoblastogenic genes expression.
MATERIALS AND METHODS
1. Reagents
Remifentanil was obtained from GlaxoSmithKline Pharmaceuticals (Rockville, MD, USA). The C2C12 cells (mouse pluripotent mesenchymal cell line) were purchased from American Type Culture Collection (ATCC; Manassas, VA, USA). Leukocyte Alkaline Phosphatase Kit (for ALP staining) was purchased from Sigma-Aldrich (St. Louis, MO). Anti-Runx2 antibodies were purchased from MBL (Nagoya, Japan), anti-osterix antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). All other chemicals and reagents were purchased from Sigma-Aldrich.
2. Cell culture and osteogenic differentiation
The C2C12 cells were maintained in Dulbecco's modified Eagle medium (DMEM; Gibco-BRL, Gaithersburg, MD, USA), containing 10% fetal bovine serum (FBS) and 1% antibiotics (50 U/ml of penicillin-G and 0.05 mg/ml of streptomycin) (Gibco-BRL), at 37℃ in a humidified 5% CO2 atmosphere. For the experiments, C2C12 cells were exposed to indicated dose of remifentanil. We assigned the group received no chemical treatment as a control group. For osteogenic differentiation of C2C12 cells, a mineralizing medium was used, which comprised of DMEM and 100 µg/ml bone morphogenic protein (BMP)-2. C2C12 cells processed with osteogenic differentiation were used as preosteoblasts. Media were changed every 48 h.
3. Cytotoxicity and cell proliferation assays
The effects of remifentanil on cell viability and proliferation were measured using the well-established colorimetric 3-(4,5-dimethylthiazol)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich) assay. In brief, the C2C12 cells were plated in 96-well plates and treated with the indicated doses of remifentanil (0, 0.1, 1, 10, and 100 ng/ml) for up to 3 days. At the end of the culture period, cells were incubated with fresh medium containing 0.5 mg/ml MTT solution for over 4 h. After incubation, the formation of blue formazan product was measured using a microplate reader at a wavelength of 570 nm.
4. Alkaline phosphatase (ALP) staining
The C2C12 cells were seeded into 48-well culture plates at a density of 2×104 cells/well and then incubated for 24 h. Cells were treated with the indicated dose of remifentanil (0, 1, and 10 ng/ml) and 100 µg/ml BMP-2, then cultured for an additional 24 and 72 h. The mineralizing medium was replaced every 48 h. To examine the ALP activity and quantitative analysis of ALP-positive area, ALP staining kit (86R-1KT, Sigma-Aldrich, St. Louis, MO, USA) was used according to the manufacturer's instructions. Sodium nitrate solution, 0.1 ml, and FRV-alkaline solution, 0.1 ml, were mixed and incubated for 2 min at room temperature. The mixed solution was diluted with 4.7 ml of double distilled water and supplemented with 0.1 ml of naphthol AS-BI alkaline solution, resulting in an alkaline-dye mixture. The cultured cells were washed with cold phosphate buffered saline (PBS) and fixed with ALP fixing solution for 30 s at room temperature. After the fixation procedure, the cells were washed three times with distilled water. The alkaline-dye mixture was added to the culture plates at a quantity of 200 µl/well. The plate was wrapped with foil and incubated for 5 to 20 min at 37℃ in the dark.
5. Boyden chamber migration assay
To examine the cell migration capacity by chemoattractant, we conducted Boyden chamber migration assay. The C2C12 cells were suspended in DMEM medium at a concentration of 1×105 cells/100 µl, and were added to the upper chamber of the transwell plates with 8 µm porosity polycarbonate membrane. Remifentanil (10 and 50 ng/ml) in DMEM medium was added to the lower chamber of each well as a chemoattractant. The C2C12 cells that migrated through the filter onto the lower side were fixed by carefully immersing the filter into methanol for 1 min. The cells were then stained with crystal violet and counted in three random microscopic fields of view per well. Each experiment was performed in duplicate and three separate experiments were performed in each group.
6. Reverse transcriptase polymerase-chain reaction (RT-PCR) analysis
Quantification of mRNA expression by PCR analysis was performed following a standard procedure. Total RNA was purified with TRIzol reagent (Invitrogen, CA, USA), and 1.5 µg of each RNA sample was reverse-transcribed using Superscript II (Invitrogen) according to the manufacturer's instructions. The primer sets used in the PCR were as follows; Runx2, 5′- CCG CAC GAC AAC CGC ACC AT-3′ (forward) and 5′- TCA GCA CAT AGC CCA CAC CG-3′ (reverse); OPN, 5′- CAG CTC AGA GGA GAA GAA GC -3′ (forward) and 5′-TGT AGG GAC GAT TGG AGT GA -3′ (reverse); BSP, 5′- TTT GCG GAA ATC ACT CTG GG -3′ (forward) and 5′- TAC GAA CAA ACA GGC AAC GA -3′ (reverse); COL1A1, 5′- GGG CAA GAC AGT CAT CGA AT -3′ (forward) and 5′-ATC ATA AGC CAA GTG GGC AG -3′ (reverse); osteocalcin, 5′- TCT TTC TGC TCA CTC TGC TG-3′ (forward) and 5′- CAG AGT TTG GCT TTA GGG CA-3′ (reverse); osterix, 5′- AAT AGT GGG CAA CTG GAG GG -3′ (forward) and 5′-GAA CAG AGC AGG CAG GTG AA -3′ (reverse); Actin, 5′- CGA TGC CCT GAG GCT CTT TT-3′ (forward) and 5′- GGG CCG GAC TCA TCG TAC TC-3′ (reverse).
7. Western blot analysis
Western blotting was conducted following a standard procedure. Briefly, C2C12 cells were disrupted in a RIPA lysis buffer (50 mM Tris; pH 8.0, 150 mM NaCl, 0.5% sodium deoxycholate, 1.5 mM MgCl2, 1 mM EGTA, 1% Triton X-100, 10 mM NaF, and complete protease inhibitor cocktail). The protein concentrations of cell lysates were determined using a detergent-compatible protein assay kit (Bio-Rad Laboratories, CA, USA), and 30–45 µg of total protein was resolved using 8–10% SDS-polyacrylamide gel electrophoresis. Separated proteins were transferred onto nitrocellulose membranes, and the membranes were blocked with 5% skim milk for 1 h. After incubation with appropriate primary/secondary antibodies, the immunoreactivity of the membranes was visualized with enhanced chemiluminescence (Amersham Pharmacia Biotech, Buckinghamshire, United Kingdom).
8. Statistical analysis
Data are presented as the mean ± standard deviation (SD) of results from three independent experiments. The Student's t-test was used to determine the significance of differences between two groups. Differences with a p value < 0.05 were regarded as being statistically significant and denoted using an asterisk.
RESULTS
1. Remifentanil did not exert cytotoxic effects or alter cell proliferation in C2C12 cells
The effect of remifentanil on the cell viability of C2C12 cells was examined using the MTT assay. As shown in Fig. 1A, the MTT assay results indicated that the viability of C2C12 cells was not affected by various concentrations of remifentanil (0–100 ng/ml). The proliferation of C2C12 cells increased with time, but there were no significant differences observed with increasing doses of remifentanil (Fig. 1B).
2. Remifentanil increased osteoblast differentiation of preosteoblastic C2C12 cells
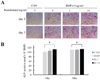
To examine the role of remifentanil in osteoblast differentiation, C2C12 cells were cultured in osteogenic media containing BMP-2 and treated with remifentanil (0, 1, and 10 ng/ml). After 24 and 72 h, the cultured C2C12 cells were stained for ALP. As shown in Fig. 2A and B, 10 ng/ml of remifentanil significantly increased ALP staining and the ALP-positive area of C2C12 cells compared to BMP-2 group (Fig. 2A, B). This result shows that remifentanil promoted osteoblast differentiation in the preosteoblastic C2C12 cells.
3. Remifentanil increased cell migration of C2C12 cells in the Boyden chamber migration assay
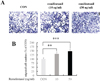
The migration of C2C12 cells was evaluated using the Boyden chamber migration assay. As shown in Fig. 3A, remifentanil increased migration of C2C12 cells compared to control. In addition, migrated cell numbers were significantly increased after transfection with 10 and 50 ng/ml of remifentanil compared to the control (Fig. 3B).
4. Remifentanil upregulated the expression of Runx2 and osterix
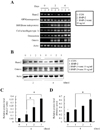
We demonstrated that remifentanil increased osteoblast differentiation (Fig. 2). To establish the mechanism involved in the effect of remifentanil on osteoblast differentiation, we investigated the mRNA expression of various osteoblastic gene markers (Runx2, OPN, BSP, COL1A1, osteocalcin, and osterix) using RT-PCR. As shown in Fig 4A, remifentanil treatment increased mRNA expression of Runx2 and osterix in the preosteoblastic C2C12 cells, compared to that in the BMP-2 group.
The protein expression of Runx2 and osterix were evaluated in C2C12 cells using western blot analysis after remifentanil treatment for 4 days. The protein level of Runx2 significantly increased after treatment with both 10 and 50 ng/ml of remifentanil, compared to the BMP-2 group (Fig. 4B, C). Remifentanil treatment increased the protein expression of Runx2 in a dose dependent manner. As shown in Fig. 4D, the protein expression of osterix significantly increased after 50 ng/ml of remifentanil compared to the BMP-2 group. These results imply that remifentanil promotes osteoblast differentiation in preosteoblastic C2C12 cells via upregulation of Runx2/osterix expression.
DISCUSSION
The primary role of osteoblasts is in the formation of bone. Besides this primary function, osteoblasts have an important role in osteoclast differentiation via cell-cell communication [11]. This communication can occur through direct contact or diffusible paracrine factors, such as growth factors, cytokines, chemokines, and other small molecules [12]. Due to their multifunction property, osteoblasts are regarded as critical cells in the overall processes of bone remodeling. We used C2C12 cells as a preosteoblast to investigate osteoblast differentiation, migration, and proliferation. C2C12 cells are a mouse pluripotent mesenchymal cell line, which are capable of rapid proliferation and myoblastic differentiation. These cells are commonly used to study the differentiation of myoblasts, osteoblasts, and myogenesis to express various target proteins and to explore mechanistic biochemical pathways in vitro [13]. In our study, C2C12 cells were suitable for the evaluation of osteoblast differentiation and for studying the expression of various osteoblastic differentiation markers by treatment with BMP-2, which causes a shift from myoblastic differentiation to osteoblastic differentiation in C2C12 cells.
In the present study, we investigated the effects of remifentanil on osteoblast differentiation by measuring the expression of osteoblast-specific genes under BMP-2 treatment. BMP has a major role in the regulation of osteoblast lineage-specific differentiation and later bone formation [14]. Among the BMPs, BMP-2, 6, and 9 have all been reported to have an important role in the induction of mesenchymal stem cell differentiation into osteoblasts [15]. BMP-2 promotes osteoblastogenesis via two types of serine/threonine kinase receptors: BMP-2 binds to the type II receptor and subsequently activates the type I receptor. Signals from the activated type I receptor are transmitted to the nucleus through the SMAD and mitogen-activated protein kinase (MAPK) pathways, which upregulate the expression of Runx2 and osterix, and facilitate osteoblast differentiation and bone formation [16]. As shown in Fig. 2, the intensity of ALP staining and the size of the ALP positive area increased after BMP-2 treatment. In addition, the mRNA expression of Runx2, BSP, COL1A1, osteocalcin and osterix were enhanced by BMP-2 treatment. These results support the idea that BMP-2 promotes osteoblastogenesis and are in accord with previous studies.
During bone remodeling, preosteoblasts or their precursors migrate into bone resorption cavities and start to form bone by filling in the bone cavities [17]. Therefore, preosteoblastic cell migration is a critical process in bone remodeling to preserve bone mass [18]. In addition, it is crucial for the repair of bone damage caused by pathologic states, such as osteoporosis and bone fracture. Preosteoblasts and osteoblasts have been reported to migrate toward chemo-attractant, but the key molecules involved in osteoblastic cell migration have not yet been studied [19]. In the present study, we have demonstrated that remifentanil increases cell migration of preosteoblasts, suggesting that remifentanil can contribute to bone healing by inducing preosteoblastic cell migration.
This study has shown that remifentanil increases osteoblastic differentiation by upregulating the expression of two key osteogenic transcription factors, Runx2 and osterix. Runx2 is an essential transcription factor that gives rise to the osteoblast lineage from mesenchymal stem cells, and once Runx2 is activated, the cells are defined as pre-osteoblasts in the osteoblast lineage [7]. Osterix is a zinc-finger transcription factor and has been shown to be required for bone formation and mineralization in vivo. In addition, osterix is a known downstream target of Runx2 during osteoblast differentiation [20]. Y. Nishio et al. [21] reported in their study that Runx2 binds to the osterix promoter and leads to the activation of osterix transcriptional regulation. Runx2 and osterix have been reported to interact and cooperatively regulate the expression of bone-specific genes. Therefore, the upregulation of Runx2 is usually accompanied by the activation of osterix.
In conclusion, remifentanil increased osteoblast differentiation in vitro through the upregulation of Runx2 and osterix expression. In addition, preosteoblastic cell migration was enhanced by remifentanil treatment. Although this study has some limitations with respect to being an in vitro study, and further studies are needed to fully elucidate our findings, it does suggest that remifentanil has the potential for assisting with bone formation and bone healing.

 XML Download
XML Download