

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Human astrocytes do not have only a lot of structural and functional properties in central nervous system (CNS) but also they are active cellular components that may response to the many specific or pathologic CNS situations and diseases (TABLE 1). Astrocytes involve in regulation of the blood-CNS interface via interactions with endothelial cells and pericytes1141526) and perform diverse supportive roles to maintain adequate neuro-metabolic microenvironment: regulation of extracellular ionic composition, pH, and maintenance of extracellular electrophysiological states and synaptic neurotransmitters activities.411131516212829)

TABLE 1
Structural and functional properties of astrocyte
![]()
Astrocytes become activated in response to many different CNS pathologies, such as stroke, trauma, infection, growth of a tumor, inflammatory — demyelinating diseases or neurodegenerative disease.2324) Instead of forming fibrous scars by fibroblast with collagen deposition in other body areas, reactive astrocytes form a glial scars in the CNS.
Numerous studies based on primary astrocyte cultures have provided a lot of information to reveal the functions of astrocytes, they are a kind of good in vitro models for studying astrocytes in themselves.15) Meanwhile, transcriptomic approach especially with human primary astrocyte (HPA) might not be only relatively rare and novel study but also could give important and essential informative clues that related with temporal activity and characteristic roles of astrocyte.
This study is about transcriptomic profiling to understand the roles of astrocytes. Elevated expression level of specific mRNAs in digital gene expression (DGE) analysis of HPA means up-regulation of specific pathway that is correlated with the major functions of human astrocyte, and vice versa.
Go to : 
MATERIALS AND METHODS
Human primary astrocyte
For this study, primary culture of astrocytes was established from embryonic human brains of 12–15 weeks' gestation. The permission to use embryonic tissues was granted by the Clinical Screening Committee for Research Involving Human Subjects of the University of British Columbia.22) Approval number of Institutional Review Board is 10-85.
HB1.F3 human neural stem cell line
The existence of immature pluripotent neural stem cells (NSCs) has been identified in the embryonic and adult mammalian brain tissue. NSCs have capability of self-proliferation and consequent differentiation into three major CNS cell types, neuron, astrocyte, and liogodendrocytes.171819)
HB1.F3 is immortalized cell line of human NSCs prepared from gestational week 14 fetal brain, which was immortalized by infection in primary culture with a retroviral vector carrying v-myc oncogene. HB1.F3 cells were grown and maintained in Dulbecco's modified Eagle's medium with high glucose (HyClone, Logan, UT, USA) containing 10% fetal bovine serum (HyClone) and 25 mg/mL gentamycin (Sigma-Aldrich, St. Louis, MO, USA).20)
Comparative analysis of digital gene expression between HPA and NSC
Library preparation and sequencing
The DGE libraries were constructed from human astrocyte tissue and HB1.F3 cell-line according to the Illumina DGE-tag profiling with NlaIII protocol (Illumina, San Diego, CA, USA). Briefly, Poly-A mRNA was isolated from 1–2 μg of total RNA using oligo-dT magnetic beads. mRNA bound to the beads was then used as a template for first strand cDNA synthesis and the second strand cDNA using random primers. The double stranded cDNA attached to oligo-dT beads was digested with NlaIII. The fragments that remained attached to the beads were ligated to the Illumina GEX NlaIII Adapter 1. Digestion with MmeI yielded the adapter tag linked to 17 base pairs (bp) of cDNA which was ligated to GEX Adapter 2 at the site of MmeI cleavage. The adaptor-ligated fragments were amplified using polymerase chain reaction and purified on 6% Novex TBE PAGE gels. A band corresponding to approximately 85 bp was cut from the gel, eluted, and concentrated by precipitation. The final library was confirmed using the Experion (Bio-Rad, Hercules, CA, USA). Sequencing was carried out using the Illumina Genome Analyzers I (Illumina).
Bioinformatics analysis
The last 9 bp sequences were trimmed out from 26 bp tags sequenced then unique tags were counted. The sequenced tags were matched with ‘top hit’ set of tags which contains best possible annotation for all possible GATC + 17bp tags in human genome/transcriptome (supported from Illumina). Differentially expressed genes were identified with no replicates method using edgeR Bioconductor software package.232) Biological pathway analysis was done with Ingenuity pathway analysis (IPA®; Ingenuity Systems, Redwood City, CA, USA) software (FIGURE 1).
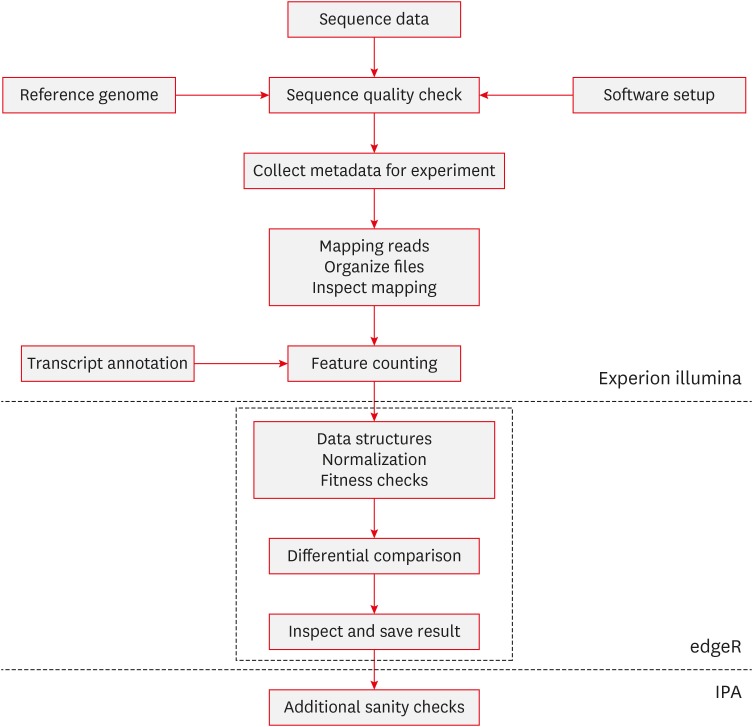
Go to :
RESULTS
The total numbers of sequence read from HPA and HB1.F3 libraries were 1,350,032 and 894,692 respectively and the status of the differentially expressed tags between HPA and HB1.F3 were checked with edgeR package. In result, 3,138 significant tags were detected, and 1,853 genes were counted among them after duplicate tags were filtered and then, up-regulated tags were 910 and down-regulated tags were 994. The Ingenuity top canonical pathways are analyzed with both up and down-regulated genes (p-value≤0.05) using IPA software and extraordinarily up or down-regulated pathways were excluded (FIGURE 2).
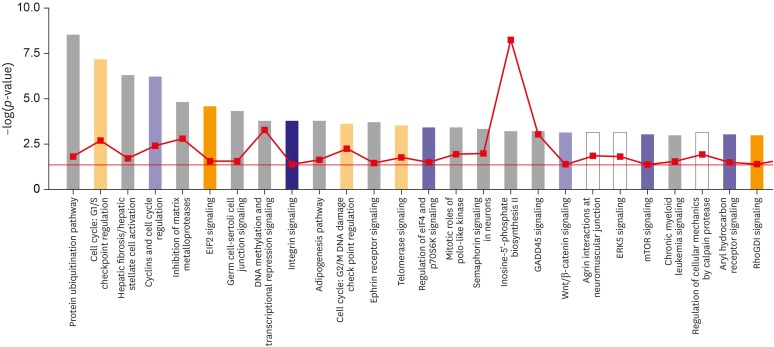 | FIGURE 2The top canonical pathways which are analyzed with both up and down-regulated genes (p-value≤0.05) using Ingenuity pathway analysis (IPA®; Ingenuity Systems, Redwood City, USA) software. Blue bars mean up-regulated pathways in astrocytes (z-score≤−2.076); orange ones mean down-regulated pathways in astrocytes (z-score≥1.5553); the activity patterns are not confirmed in gray-colored pathways. Orange pints are the ratio of the number of genes that meet the cut off criteria/the number of genes that make up that pathway.
|
Up-regulated pathways in HPA
Ingenuity top canonical pathways that are up-regulated in HPA as compared with HB1.F3 cell line were ‘Cyclins and cell cycle regulation,’ ‘Integrin signaling,’ ‘Regulation of eukaryotic translation initiation factor 4 (eIF4) and p70S6K signaling,’ ‘Wnt/β-catenin signaling,’ ‘mammalian the target of rapamycin (mTOR) signaling,’ ‘Aryl hydrocarbon receptor signaling,’ ‘Hippo signaling,’ ‘RhoA signaling,’ ‘Signaling by Rho family GTPases,’ and ‘Glioma signaling’ pathways (TABLE 2).
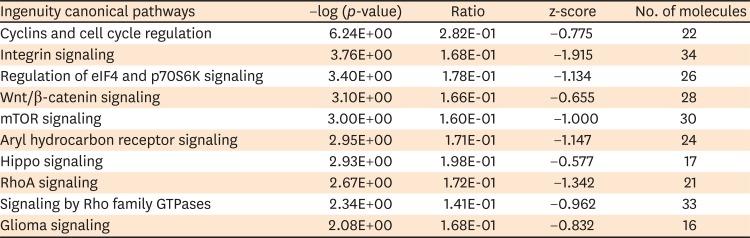
TABLE 2
Ingenuity top canonical pathways up-regulated in human primary astrocyte
![]()
Down-regulated pathways in HPA
Ingenuity top canonical pathways that are down-regulated in HPA as compared with HB1.F3 cell line were ‘Cell cycle: G1/S checkpoint regulation,’ ‘Eukaryotic translation initiation factor-2 (eIF2) signaling,’ ‘Cell cycle: G2/M DNA damage checkpoint regulation,’ ‘Telomerase signaling,’ ‘RhoGDI signaling,’ ‘NRF2-mediated oxidative stress response,’ ‘Extracellular-regulated kinase (ERK)/mitogen activated protein kinase (MAPK) signaling,’ ‘Ataxia telangiectasia mutated (ATM) signaling,’ ‘Pancreatic adenocarcinoma signaling,’ ‘Vascular endothelial growth factor (VEGF) signaling, and ‘Role of Csk homologous kinase (CHK) proteins in cell cycle checkpoint control’ pathways (TABLE 3).
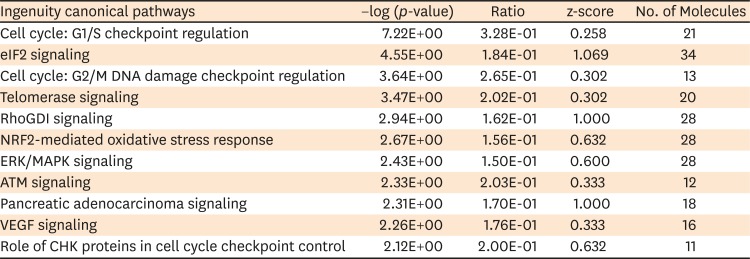
TABLE 3
Ingenuity top canonical pathways down-regulated in human primary astrocyte
eIF2: eukaryotic translation initiation factor-2, ERK: extracellular-regulated kinase, MAPK: mitogen activated protein kinase, ATM: ataxia telangiectasia mutated, VEGF: vascular endothelial growth factor, CHK: homologous kinase.
![]()
Go to :
DISCUSSION
Large amount of diverse RNAs play many important roles in mediating from DNAs to synthesized protein and are influenced by various regulatory mechanisms. The pattern of RNA populations might reflect the state and circumstances of each characteristic cells. The measurement of mRNA populations in a cell group of interest is called gene expression profiling or transcript profiles. Many popular high-throughput gene analysis techniques such as quantitative real-time polymerase chain reaction, serial analysis of gene expression (SAGE), and DNA microarray used to be applied for transcriptome profiling to understand the molecular background of specific cells in various diseases. But these techniques are not of great utility regarding feasibility, reproducibility, cost and the amount of material required for analysis.
In comparison to hybridization-based methods such as microarrays, direct sequencing methods have the advantages to detect variation of transcriptomes and to discover novel small RNAs. Despite various advantages over hybridization methods, the use of sequencing-based methods has been hindered mostly by the expense of capillary DNA sequencing. In contrast, sequence tag based approaches like SAGE and massively parallel signature sequencing can measure the expression of both known and unknown genes. With the advent of massively parallel sequencing platforms, the cost and labor involved in sequencing have been significantly reduced especially for global gene expression pattern analysis.527)
In this study, the cell cycle related pathways such as ‘Cell cycle: G1/S checkpoint regulation,’ ‘G2/M DNA damage checkpoint regulation’ and ‘Role of CHK proteins in cell cycle checkpoint control’ are generally down-regulated except only ‘Cyclin and cell cycle regulation’ pathway which is up-regulated in contrast. Because astrocytes are fully differentiated cells with individual cellular activities in the specific areas in CNS, and could be activated properly in case of specific abnormal or pathologic circumstance, the study results stand to reason.
Integrins are cell surface glycoproteins that are involved in cell-to-cell and cell-to-extracellular matrix interactions. They have important roles in supporting cytoskeleton, anchorage-dependent proliferation, and signal transduction. These interactions are deeply related with the known functions of astrocyte.10) Therefore, it is the reason why ‘Integrin Signaling’ is up-regulated in HPA.
The Wnt that is derived from ‘Wingless,’ the Drosophila melanogaster segment-polarity gene, and the vertebrate homologue Integrase-1. The Wnt proteins are a kind of extracellular growth factors and the signaling pathways essential roles in embryonal development, tissue regeneration, and many other processes. In CNS it is known to regulate the balance between self-renewal and differentiation,31) especially it increases in proliferating astrocytes during post-traumatic gliogenesis in the adult brain and it is essential for development of astrocyte.7) Meanwhile, Hippo pathway has been known to promote cell death, differentiation, and inhibit cell proliferation. It has the functions to control organ size, therefore, its dysfunction can cause tumor or organ degeneration.30) Therefore, up-regulated appearance of Wnt and Hippo signaling pathway in comparison with HB1.F3 is very natural and important with characteristic functions of astrocytes.
mTOR pathway is a central regulator of glial and glioma cell growth and it involves in essential role in protein homeostasis in CNS. It also is related to neurodegenerative diseases such as Alzheimer's, Parkinson's by aberrant accumulation of misfolded proteins leading neuronal loss and thus relative up-regulation of mTOR pathway is a reflection of the functional activities of astrocytes.38) eIF4 and p70S6K play important roles in translational regulation. Recruitment of mRNAs to ribosomes to initiate translation is mediated by initiation factors of the eIF4 group and poly(A)-binding protein.6) Both mTOR and regulation of eIF4 and p70S6K signaling pathway might be involved in protein synthesis in the astrocytes and up-regulated. eIF2 is a GTP-binding protein that escorts the initiation specific form of met-tRNA onto the ribosome as a role in identifying the translational start site. eIF2 pathway was down-regulated in HPA in contrast with eIF4.
The GTPase family of small GTP-binding proteins comprises a group of signaling molecules that are activated by diverse factors or molecules. They regulate a wide range of cellular biological activities. The mammalian GTPase family currently consists of three subgroups: Rho, Rac and Cdc42. Each controls the formation of a distinct cytoskeletal element in the cells. Rho regulates bundling of actin filaments into stress fibers and the formation of focal adhesion complexes. The small GTPases act as molecular switches, cycling between an active GTP-bound state and an inactive GDP-bound state. The dynamic organization of the actin cytoskeleton provides the force for cell motility Rho family proteins regulate a wide spectrum of cellular functions such as cytoskeletal organization, membrane transporting, cell proliferation, cell motility and transcriptional regulation.12) On the other hand, Rho GDI signaling pathway negatively regulate Rho family GTPases. The inhibitory activity of GDI derives both from an ability to bind the carboxy-terminal isoprene of Rho GTPases and extract them from membranes, and from inhibition of GTPase cycling between the GTP- and GDP-bound states.9) In this study, the ‘Signaling by Rho family GTPases’ pathway was up-regulated and whereas, Rho GDI signaling pathway was down-regulated. These diverse activities of Rho family related pathways might have intimate correlations of known variable function of astrocytes.
The ERK/MAPK pathway is a key pathway that transduces cellular information on cell division,25) the ATM signaling pathway is respond to DNA stand breaks during cell proliferation, and the VEGF signaling pathway's main role is angiogenesis during embryonal development. So, these pathways are naturally down-regulated in mature astrocytes. Because the telomere is elongated in HB1.F3 cell line but the telomere is maintained in HPA, the ‘Telomerase signaling’ pathway activity was detected as relatively down regulated in HPA.
In this study, HPA and HB1.F3 are not harvested in one same individual human being. So, its bias exists. If we harvested HPA and HB1.F3 in one fetus, this study could be ideal. On the other hand, if the brain tissues are harvested in different regions, they may not show the common epigenetic patterns due to the differences of functions which are recommend typically in each brain areas even though the cell components are same as astrocytes. Therefore, additional further studies with well-designed methods are needed on this.
Go to :
CONCLUSION
These relatively up-regulated or down-regulated characteristic pathways have very intimate correlation with proper activities of astrocyte in specific CNS circumstance, therefore this study would be a good reference to understand the astrocyte function at molecular level and to develop the test of DGE pattern analysis of astrocyte as new powerful clinical tools in many CNS disease in the future.
Go to :
 XML Download
XML Download