

 PDF
PDF Citation
Citation Print
Print



Abbreviations
BMDM
bone marrow-derived macrophage
cDNA
complementary DNA
DMEM
Dulbecco's modified Eagle's medium
EKSA
ethanol-killed Staphylococcus aureus
HDAC
histone deacetylase
iNOS
inducible nitric oxide synthase
LB
Luria-Bertani
LTA
lipoteichoic acid
MPN
mepenzolate bromide
MTT
2,5-diphenyl-2H-tetrazolium bromide
NO
nitric oxide
PAMP
pathogen-associated molecular pattern
P-STAT1
phosphorylated STAT1
PTX
pertussis toxin
Sa.LPP
Staphylococcus aureus lipoprotein
SAHA
suberoylanilide hydroxamic acid
SCFA
short-chain fatty acid
TBS
Tris-buffered saline
TBST
TBS containing 0.05% Tween 20
TSA
trichostatin A
INTRODUCTION
Staphylococcus aureus is a Gram-positive pathogenic bacterium that can cause severe inflammation leading to septic shock and inflammatory bowel diseases (12). TLR2 ligands of Gram-positive bacteria (including S. aureus) are responsible for inducing severe inflammation in the host cell (3). In S. aureus, lipoproteins and lipoteichoic acid (LTA) are well known TLR2 ligands responsible for inducing nitric oxide (NO) and/or proinflammatory cytokine production in macrophages (45). Many recent studies have suggested that the bacterial lipoprotein is a potent immunostimulator in macrophages (67). Wild-type and LTA-deficient, but not lipoprotein-deficient, S. aureus induces NO production in macrophages (8). In addition, lipoprotein-deficient Streptococcus gordonii is less effective in inducing NO production than wild-type or LTA-deficient S. gordonii in macrophages (6). Furthermore, wild-type, but not lipoprotein-deficient, S. aureus potently induces IL-8 induction in human intestinal epithelial cells (5) and osteoclast activation (9).
NO is a small molecule that can regulate a variety of physiological functions such as innate immune responses, vascular homeostasis, and neurotransmission (10). In mammalian cells, inducible NO synthase (iNOS) can induce a micromolar level of NO by immune cell activation, which can evoke septic shock, autoimmune diseases, and chronic inflammatory diseases (11). Excessive NO production by iNOS is observed in patients with septic shock or inflammatory bowel diseases (1213). NF-κB activation and type I IFN-mediated STAT1 phosphorylation are essential for iNOS expression in macrophages (14). In S. aureus, lipoprotein is the major cell wall component for inducing excessive NO production through TLR2-mediated iNOS induction in macrophages (8). Even though NO production induced by S. aureus lipoprotein (Sa.LPP) is known to be detrimental to the host, little is known about molecules that could potentially inhibit excessive inflammation.
Short-chain fatty acids (SCFAs) are metabolites produced by intestinal microbiota through fermentation of undigested carbohydrates and dietary fibers (15). Butyrate, propionate, and acetate are the predominant forms of SCFAs, which have anti-inflammatory properties (1617). Butyrate has beneficial roles by having anti-inflammatory effects on diseases such as inflammatory bowel disease or sepsis (1819). Furthermore, SCFAs regulate immune cell differentiation and function through the inhibition of histone deacetylase (HDAC) and activation of G protein-coupled receptors (2021). SCFAs also downregulate NO production by IFN-γ through the inhibition of NF-κB and ERK signaling in macrophages (22). Although SCFAs have been suggested as anti-inflammatory molecules (2324), it is not fully understood whether SCFAs regulate bacterial lipoprotein-mediated NO production in macrophages. In this study, we investigated whether SCFAs inhibit Sa.LPP-induced NO production in macrophages.
MATERIALS AND METHODS
Bacteria, reagents, and chemicals
S. aureus RN4220 was kindly provided by Prof. Bok Luel Lee (Pusan National University, Busan, Korea). Luria-Bertani (LB) broth was purchased from LPS Solution (Daejeon, Korea). Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were purchased from Welgene (Gyeongsan, Korea) and Gibco (Burlington, ON, Canada), respectively. Recombinant murine M-CSF was obtained from CreaGene (Seongnam, Korea). Sodium acetate, sodium propionate, sodium butyrate, trichostatin A (TSA), suberoylanilide hydroxamic acid (SAHA), mepenzolate bromide (MPN), pertussis toxin (PTX), Triton X-114, octyl β-D-glucopyranoside, and thiazolyl blue tetrazolium bromide were purchased from Sigma-Aldrich Inc. (St. Louis, MO, USA). Anti-iNOS rabbit polyclonal IgG antibody was obtained from Upstate Biotechnology (Lake Placid, NY, USA). Anti-acetyl-histone H3 (Lys9) polyclonal antibody was purchased from Millipore (Billerica, MA, USA). Anti-STAT1 and -phosphorylated STAT1 (P-STAT1) rabbit polyclonal antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-β-actin mouse monoclonal antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All other reagents were purchased from Sigma-Aldrich Inc. unless indicated otherwise.
Preparation of ethanol-killed S. aureus (EKSA)
Techniques used to prepare EKSA were previously described (25). Briefly, S. aureus was cultured in LB medium at 37°C to mid-log phase. The bacterial pellet was collected, incubated, and shaken with 70% ethanol in PBS at room temperature for 2 h. After washing twice with PBS, bacterial killing was confirmed by spreading on an LB-agar plate at 37°C for 48 h. No bacterial colonies were observed.
Culture of RAW 264.7 cells
RAW 264.7 (TIB-71) was obtained from the American Type Culture Collection (Manassas, VA, USA). The cells were cultured in DMEM supplemented with 10% FBS, penicillin (100 U/ml) and streptomycin (100 μg/ml) at 37°C in a humidified incubator with 5% CO2.
Preparation of bone marrow-derived macrophages (BMDMs)
Animal experiments were conducted under the approval of the Institutional Animal Care and Use Committee of Seoul National University (SNU-170103-3). C57BL/6 mice were purchased from Orient Bio (Seongnam, Korea). Bone marrow cells were prepared from 8-week-old mice as previously described (26). Bone marrow cells were differentiated into BMDMs with DMEM supplemented with 10% FBS, penicillin (100 U/ml), streptomycin (100 μg/ml) and M-CSF (20 ng/ml) for 5 days. BMDMs (5×105 cells/ml) were stimulated with the indicated stimuli for 24 h.
Measurement of NO production
RAW 264.7 cells (3×105 cells/ml) were stimulated with the indicated stimuli for 24 h. Nitrite in cell culture supernatants was measured to determine NO as previously described (27). Briefly, an equal volume of Griess reagent (1% sulfanilamide, 0.1% naphthylethylenediamine dihydrochloride, and 2% phosphoric acid) was added to culture supernatants and incubated at room temperature for 5 min. Then, the concentration of nitrite was measured at 540 nm with a microtiter plate reader (Versamax, Molecular Devices Corporation, Sunnyvale, CA, USA) using NaNO2 as a standard.
Cell viability assay
RAW 264.7 cells (3×105 cells/ml) were stimulated with the indicated stimuli for 24 h. Then, the cells were treated with 0.5 mg/ml of 2,5-diphenyl-2H-tetrazolium bromide (MTT) reagent for 2 h. The precipitate (formazan) was dissolved with dimethyl sulfoxide and the absorbance was measured at 570 nm with a microtiter plate reader.
Western blotting
RAW 264.7 cells (3×105 cells/ml) were treated with the indicated stimuli for various time periods. After washing with PBS, the cells were lysed in a lysis buffer (50 mM Tris-HCl, 1% NP-40, 0.25% sodium deoxycholate, 1 mM EDTA, and 150 mM NaCl) containing protease/phosphatase inhibitors (1 mM PMSF, 1 μg/ml aprotinin, 1 μg/ml leupeptin, 1 mM Na3VO4, and 1 mM NaF). The lysate was separated by SDS-PAGE and transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% bovine serum albumin or 5% skim milk in Tris-buffered saline (TBS) at room temperature for 1 h and incubated overnight with specific antibodies against iNOS, H3K9Ac, P-STAT1, STAT1, or β-actin at 4°C. After incubation, membranes were washed three times with TBS containing 0.05% Tween 20 (TBST) and incubated with HRP-conjugated secondary antibody (Southern Biotechnology, Birmingham, AL, USA) at room temperature for 1 h. Membranes were rinsed 3 times with TBST and developed with SUPEX ECL solution (Neuronex, Pohang, Korea) as previously described (28).
RT-PCR
RAW 264.7 cells (3×105 cells/ml) were treated with the indicated stimuli for various time periods. Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instruction. Complementary DNA (cDNA) was synthesized from total RNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA) and random hexamers (Roche, Basel, Switzerland). PCR was performed as previously described (29). Briefly, cDNA amplification was performed with EmeraldAmp PCR Master Mix (Takara Biomedical Inc., Osaka, Japan) and primers specific to murine iNOS (forward primer: 5′-GGA TAG GCA GAG ATT GGA GG-3′, reverse primer: 5′-AAT GAG GAT GCA AGG CTG G-3′), murine IFN-β (forward primer: 5′-AAT TCT CCA GCA CTG GGT GG-3′, reverse primer: 5′-CCA GGC GTA GCT GTT GTA CT-3′), or β-actin (forward primer: 5′-GTG GGC CCC AGG ACC A-3′, reverse primer: 5′-CTC CTT AAT GTC ACG CAC GAT TTC-3′).
Transient transfection and reporter gene assay
RAW 264.7 cells (1×106 cells/ml) were seeded on 6-well plate at 3 h before transfection. The cells were transfected with pNF-κB-Luc (Clontech, Mountain View, CA, USA) using Attractene reagent (Qiagen, Valencia, CA, USA) for 3 h according to the manufacturer's instructions. After the transfection period, the cells were harvested and plated on 96-well plates for 3 h. The cells were then stimulated with the indicated stimuli for 21 h. For reporter gene assays, the cells were lysed with Gro-lysis buffer (Promega), and the luciferase activity in supernatants was determined using Luciferase Reporter Assay System (Promega) with SPARK 10M multimode microplate reader (Tecan Group Ltd., Männedorf, Switzerland).
Preparation of lipoproteins from S. aureus
Lipoproteins were prepared from S. aureus as previously described (30). Briefly, a bacterial pellet was sonicated and vigorously mixed with 2% Triton X-114 in TBS at 4°C for 2 h. After the incubation period, debris was removed by centrifugation at 10,410 ×g for 10 min, and the supernatants containing lipoproteins were collected and mixed with an equal volume of TBS. The supernatants were incubated at 37°C for 15 min, and the lipoprotein-containing phase was obtained by centrifugation. Lipoproteins were then isolated by methanol precipitation and dissolved in 10 mM of octyl β-D-glucopyranoside in PBS.
Statistical analysis
All experiments were performed at least three times. Mean values±standard deviations were obtained from triplicate samples for each treatment group. Statistical significance was examined with ANOVA using GraphPad Prism 5 (GraphPad Software Inc., La Jolla, CA, USA). An asterisk (*) indicates treatment groups that were significantly different from the control group at p<0.05.
RESULTS
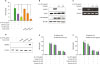
Butyrate and propionate, but not acetate, inhibit Sa.LPP-induced NO production in RAW 264.7 cells
It has been reported that SCFAs have inhibitory effects against S. aureus-induced immune responses in host cells (3132). To examine the ability of SCFAs to regulate live S. aureus- or Sa.LPP-induced NO production in macrophages, RAW 264.7 cells were stimulated with live S. aureus in the presence or absence of SCFAs for 3 h and further incubated with gentamicin to prevent bacterial growth for 21 h. Butyrate and propionate, but not acetate, remarkably inhibited live S. aureus-induced NO production (Fig. 1A). Similar to the results of live S. aureus, butyrate and propionate, but not acetate, inhibited EKSA-induced NO production (Fig. 1B). To examine if SCFAs also inhibited Sa.LPP-induced NO production, RAW 264.7 cells were stimulated with purified Sa.LPP (5 μg/ml) in the presence or absence of SCFAs. Butyrate and propionate significantly inhibited Sa.LPP-induced NO production, whereas such inhibitory effects were not observed in acetate-treated groups (Fig. 1C). Of note, butyrate potently inhibited live S. aureus-, EKSA-, or Sa.LPP-induced NO production even at 1 mM. Moreover, 3 mM of butyrate and propionate, but not acetate, significantly inhibited Sa.LPP-induced NO production in primary macrophages (Fig. 1D). To exclude the possibility that changes in cell viability had an impact on NO production, RAW 264.7 cells were stimulated with Sa.LPP in the presence or absence of SCFAs, and cell viability was determined using the MTT assay (Fig. 1E). There was no cytotoxicity even when SCFAs inhibited the Sa.LPP-induced NO production. These results suggest that butyrate and propionate inhibit Sa.LPP-induced NO production in murine macrophages.
Figure 1
Butyrate and propionate, but not acetate, inhibit S. aureus or its lipoprotein-induced NO production in murine macrophages. (A) RAW 264.7 cells (3×105 cells/ml) were treated with 1×108 CFU/ml of live S. aureus in the presence (0.3, 1, or 3 mM) or absence of acetate, propionate, or butyrate for 3 h and further incubated with 200 μg/ml of gentamicin for 21 h. (B) RAW 264.7 cells (3×105 cells/ml) were treated with 50 μg/ml of EKSA in the presence (0.3, 1, or 3 mM) or absence of acetate, propionate, or butyrate for 24 h. (C) RAW 264.7 cells (3×105 cells/ml) were stimulated with 5 μg/ml of Sa.LPP in the presence (0.3, 1, or 3 mM) or absence of acetate, propionate, or butyrate for 24 h. Nitrite in the culture supernatant was measured to determine NO concentration using the Griess reagent. (D) BMDMs (5×105 cells/ml) were stimulated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 24 h, and nitrite was measured. (E) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP and cultured in the presence (0.3, 1, or 3 mM) or absence of acetate, propionate, or butyrate for 24 h. Cell viability was measured by MTT assay. Data are mean values±standard deviations of 3 independent experiments.
N.D., not detected.
*Asterisks indicate significant differences (p<0.05) compared with the appropriate controls.

Butyrate and propionate, but not acetate, inhibit Sa.LPP-induced iNOS expression in RAW 264.7 cells
NO is produced by iNOS, which is one of the NOS isoforms (33). Therefore, we examined the inhibitory potency of SCFAs on Sa.LPP-induced iNOS expression in RAW 264.7 cells. Butyrate and propionate, but not acetate, significantly inhibited iNOS expression at the protein level (Fig. 2A). Butyrate inhibited Sa.LPP-induced iNOS protein expression more potently than propionate. Similar results were observed at the mRNA level. Butyrate and propionate, but not acetate, reduced Sa.LPP-induced iNOS mRNA expression levels in RAW 264.7 cells (Fig. 2B). These results indicate that butyrate and propionate downregulate Sa.LPP-induced iNOS expression, which is responsible for NO production in murine macrophages.
Figure 2
Butyrate and propionate, but not acetate, inhibit Sa.LPP-induced iNOS expression in RAW 264.7 cells. (A) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 15 h. Cell lysates were analyzed to measure iNOS or β-actin expression by Western blotting. (B) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 12 h. Total RNA was isolated, and the mRNA expression levels of iNOS or β-actin was examined by RT-PCR. Relative expression levels of iNOS protein and mRNA to those of β-actin were measured to obtain mean values±standard deviations by densitometry from three independent experiments (right panels).
*Asterisks indicate significant differences (p<0.05) between the Sa.LPP treatment group and the Sa.LPP with SCFA treatment group.
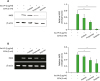
Butyrate and propionate inhibit Sa.LPP-induced NF-κB activation, STAT1 phosphorylation, and IFN-β mRNA expression in RAW 264.7 cells
NF-κB activation through TLR2 signaling and STAT1 phosphorylation through type I IFN are essential signaling pathways responsible for the transcription of iNOS (14). To determine if SCFAs regulate the Sa.LPP-mediated NF-κB activation, RAW 264.7 cells were transfected with NF-κB reporter plasmid and treated with Sa.LPP in the presence or absence of SCFAs. NF-κB transcriptional activity was analyzed by reporter gene assay. Butyrate and propionate significantly inhibited Sa.LPP-induced NF-κB activation in RAW 264.7 cells, whereas acetate did not (Fig. 3A). In addition, butyrate and propionate, but not acetate, remarkably inhibited Sa.LPP-induced phosphorylation of NF-κB p65 (Fig. 3B). To determine whether SCFAs regulate Sa.LPP-induced STAT1 phosphorylation, RAW 264.7 cells were stimulated with Sa.LPP in the presence or absence of SCFAs for 4 h. Butyrate and propionate significantly inhibited Sa.LPP-induced STAT1 phosphorylation, whereas acetate did not (Fig. 3C). Furthermore, butyrate and propionate significantly inhibited Sa.LPP-induced IFN-β expression (Fig. 3D), which is responsible for activating STAT1 through type I IFN receptors. These results suggest that butyrate and propionate, but not acetate, significantly inhibit Sa.LPP-mediated iNOS-inducing factors including NF-κB activation, STAT1 phosphorylation, and IFN-β expression in macrophages.
Figure 3
Butyrate and propionate, but not acetate, inhibit Sa.LPP-induced NF-κB activation, STAT1 phosphorylation, and IFN-β mRNA expression in RAW 264.7 cells. (A) RAW 264.7 cells (1×106 cells/ml) were transfected with a firefly luciferase reporter plasmid regulated by NF-κB for 3 h. After the cells were harvested and plated on 96-well plates for 3 h, the cells were treated with 5 μg/ml of Sa.LPP and cultured in the presence or absence of 3 mM acetate, propionate, or butyrate for additional 21 h. Cell lysates were mixed with firefly luciferase substrate to determine luciferase activity. (B) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 3 h. Cell lysates were analyzed to measure phosphorylated NF-κB p65 or β-actin by Western blotting. (C) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 3 h. Cell lysates were analyzed to measure phosphorylated STAT1, nonphosphorylated STAT1 or β-actin by Western blotting. (D) RAW 264.7 cells (5×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of 3 mM acetate, propionate, or butyrate for 6 h. Total RNA was isolated, and the mRNA expression level of IFN-β was examined by RT-PCR. Relative expression levels of P-STAT1 to STAT1 and IFN-β to β-actin were measured to obtain mean values±standard deviations by densitometry from 3 independent experiments (right panels).
*Asterisks indicate significant differences (p<0.05) between the Sa.LPP treatment group and the Sa.LPP with SCFA treatment group.
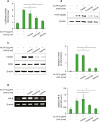
Butyrate inhibits Sa.LPP-induced NO production by inhibiting HDAC
Inhibition of HDAC and activation of G protein-coupled receptor, such as GPR43 and GPR109A, are well-known immunoregulatory mechanisms of butyrate (18). To determine if HDAC inhibition is involved in the SCFA-mediated inhibitory effects on Sa.LPP-induced NO production, RAW 264.7 cells were pretreated with an HDAC inhibitor, SAHA or TSA, for 1 h and then stimulated with Sa.LPP for an additional 23 h. SAHA and TSA inhibited Sa.LPP-induced NO production in RAW 264.7 cells, to a similar extent as butyrate and propionate (Fig. 4A). TSA remarkably inhibited Sa.LPP-induced STAT1 phosphorylation and NF-κB activation (Fig. 4B). TSA also inhibited Sa.LPP-induced IFN-β mRNA expression (Fig. 4C). To confirm that butyrate and propionate induce histone acetylation at lysine residues in the presence of Sa.LPP, RAW 264.7 cells were stimulated with Sa.LPP in the presence or absence of SCFAs for 3 h. Western blotting showed that butyrate (potently) and propionate (modestly) induced histone acetylation whereas acetate hardly showed such an effect under the same conditions (Fig. 4D). Next, to determine whether signaling through GPR43 or GPR109A is involved in the inhibitory effects of SCFAs on Sa.LPP-induced NO production, RAW 264.7 cells were pretreated with inhibitors for GPR109A (MPN) or G protein-coupled receptors (PTX) for 1 h and then stimulated with Sa.LPP and SCFAs for an additional 23 h. The inhibitory effects of butyrate and propionate in Sa.LPP-induced NO production were not affected by pretreatment of those inhibitors (Fig. 4E and F). These results suggest that HDAC inhibition, but not G protein-coupled receptor signaling, is the key molecular mechanism for the inhibitory effect of butyrate and propionate on Sa.LPP-induced NO production in murine macrophages.
Figure 4
Butyrate and propionate might inhibit Sa.LPP-induced NO production by inhibiting HDAC. (A) RAW 264.7 cells (3×105 cells/ml) were pre-treated with HDAC inhibitors (0.5 μM SAHA or 0.05 μM TSA) or SCFAs (3 mM of acetate, propionate, or butyrate) for 1 h and subsequently stimulated with 5 μg/ml of Sa.LPP for additional 23 h. Nitrite in the culture supernatant was measured to determine NO concentration using the Griess reagent. (B) RAW 264.7 cells (3×105 cells/ml) were pre-treated with 0.01 or 0.05 μM of TSA for 1 h and subsequently stimulated with 5 μg/ml of Sa.LPP for additional 3 h. Cell lysates were analyzed to measure phosphorylated STAT1, nonphosphorylated STAT1, phosphorylated NF-κB p65 or β-actin by Western blotting. (C) RAW 264.7 cells (5×105 cells/ml) were pre-treated with 0.01 or 0.05 μM of TSA for 1 h and subsequently stimulated with 5 μg/ml of Sa.LPP for an additional 6 h. Total RNA was isolated, and the mRNA expression level of IFN-β was examined by RT-PCR. (D) RAW 264.7 cells (3×105 cells/ml) were treated with 5 μg/ml of Sa.LPP in the presence or absence of acetate, propionate, or butyrate for 4 h. Cell lysates were obtained and analyzed by Western blotting to measure histone acetylation at lysine residues. (E, F) RAW 264.7 cells (3×105 cells/ml) were pre-treated with (E) 0.01 μM of MPN, or (F) 0.01 μg/ml of PTX for 1 h and subsequently stimulated with 5 μg/ml of Sa.LPP for an additional 23 h. Nitrite in the culture supernatant was measured to determine NO concentration using the Griess reagent. Data are mean values±standard deviation of 3 independent experiments.
*Asterisks indicate significant differences (p<0.05) compared with the appropriate controls.
DISCUSSION
The lipoprotein of Gram-positive bacteria is a potent stimulator that can cause severe inflammation in the host cell (34). In this study, we demonstrated that butyrate and propionate inhibited Sa.LPP-induced NO production through the downregulation of iNOS expression. Butyrate and propionate also inhibited NF-κB activation, IFN-β expression, and STAT1 phosphorylation, which are essential for the induction of iNOS gene expression. Moreover, butyrate and propionate inhibited NO production, which might be via inhibition of HDAC. Butyrate was much more potent than propionate in all of the aforementioned data. Therefore, these results suggest that SCFAs butyrate and propionate are anti-inflammatory for downregulating Sa.LPP-induced NO production in macrophages.
Butyrate seems to inhibit pathogen-associated molecular pattern (PAMP)-induced inflammatory conditions in general, considering our current study and a previous study showing that butyrate inhibits peptidoglycan-induced TNF-α and IL-1β expression in THP-1 cells (35). Butyrate also inhibits Pam3CSK4 (a synthetic lipopeptide mimicking bacterial lipoproteins)-induced IL-8 production in human intestinal epithelial cells (36). Furthermore, in dendritic cells, butyrate effectively inhibits NO, IL-12p40, and IFN-γ expression by LPS, which is one of the major PAMPs of Gram-negative bacteria (37). These results indicate that butyrate could be a potent immunosuppressive molecule in various inflammatory conditions caused by PAMPs from both Gram-positive and Gram-negative bacteria.
In general, activation of NF-κB is stronger in patients who suffer from inflammatory diseases than in healthy people (38). Lipoproteins of Gram-positive bacteria including S. aureus (8), Streptococcus pneumoniae (39), and S. gordonii (6) are major TLR2 ligands. Lipoprotein-induced TLR2 activation causes MyD88-dependent NF-κB activation, which is responsible for inducing pro-inflammatory responses in human intestinal epithelial cells (5). In this study, we found that butyrate potently inhibits Sa.LPP-induced NF-κB activation. The inhibitory effect of butyrate on NF-κB activation seems to be a general phenomenon, considering that butyrate inhibits LPS-induced transcriptional activity of NF-κB in macrophages (40) and TNF-α-induced nuclear translocation of NF-κB in human colon cells (41). Therefore, butyrate has anti-inflammatory effects against inflammation caused by bacterial infections via inhibitory action on NF-κB.
HDAC activity is required for STAT1 signaling (42) and bacterial lipoprotein efficiently induces TLR2-mediated IFN-β expression in endolysosomal compartments leading to STAT1 phosphorylation in macrophages (43). Interestingly, we showed that butyrate and propionate inhibit Sa.LPP-induced IFN-β expression and STAT1 phosphorylation. HDAC inhibition seems to be important for the inhibitory effect of butyrate and propionate, considering that HDAC inhibition can induce ubiquitination and proteasomal degradation of interferon regulatory factor 1 (44), which is an essential transcription factor for inducing TLR2/MyD88-mediated IFN-β expression in macrophages (43). Furthermore, we showed that butyrate and propionate promote histone acetylation and that the inhibition of HDAC downregulates Sa.LPP-induced IFN-β expression and STAT1 phosphorylation.
Concordant with our study, Park et al. (22) demonstrated that SCFAs (butyrate, phenylbutyrate, and phenylacetate) downregulate IFN-γ-induced NO production through the inhibition of NF-κB signaling pathways in murine macrophages. However, there are some differences between their results and our results. First, they showed that not only butyrate and phenylbutyrate, but also phenylacetate, downregulate IFN-γ-induced NO production. However, we showed that acetate does not downregulate S. aureus- or Sa.LPP-induced NO production. These results suggest that the anti-inflammatory potency of SCFAs may vary depending on the subtype of SCFAs. Second, while Park et al. (22) showed that SCFAs do not affect IFN-γ-induced Jak/STAT signaling, our current study showed that butyrate and propionate regulate STAT1 signaling. Although further studies are needed to elucidate the cause of different results, this might be at least partially due to a difference in stimuli. Notably, IFN-γ (used in their study) is a sort of a host-derived cytokine that directly activates Jak/STAT signaling leading to iNOS expression. In contrast, S. aureus and its lipoproteins (used in our study) primarily stimulate pattern-recognition receptors such as TLR2, which indirectly activates Jak/STAT signaling through type I IFN responses in macrophages (4345). Indeed, our study further demonstrated that butyrate and propionate downregulate Sa.LPP-induced IFN-β expression.
We showed that propionate also inhibits Sa.LPP-induced NO production, but its inhibitory potency is less than that of butyrate, while acetate does not inhibit production at all. Different immunomodulatory effects of SCFAs have been reported in other experimental conditions. For example, butyrate more potently inhibits LPS-induced NO, IL-6, and IL-12p40 expression than propionate or acetate in BMDMs (18). Furthermore, we have reported that only butyrate, but not propionate or acetate, potentiates oral cholera vaccine-induced CCL20 secretion in human intestinal epithelial cells (46). The different immunomodulatory potency of each SCFA might be due to differences in HDAC inhibitory activity, considering that butyrate more potently induces histone acetylation than propionate or acetate (47). The results of this study showed that butyrate more potently induces histone acetylation compared to propionate in Sa.LPP-treated RAW 264.7 cells, whereas acetate does not.
Lipoprotein is a major virulence factor in S. aureus and is responsible for inducing excessive pro-inflammatory cytokines, leading to severe sepsis in the host (4849). Therefore, identification of an anti-inflammatory molecule is important to prevent excessive inflammation from S. aureus infections. This study suggests that butyrate is a potent anti-inflammatory molecule that can inhibit lipoprotein-induced NO production. Butyrate may act as a regulator against excessive inflammation caused by bacterial infection.
 XML Download
XML Download