

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Invariant NKT (iNKT) cells are a small subset of thymus-generated αβ T lineage cells that produce large quantities of cytokines to control both innate and adaptive immunity (12). iNKT cells develop from immature CD4+CD8+ double positive (DP) thymocytes upon strong agonistic stimulation of their TCRs by glycolipid-loaded CD1d molecules (34). The exact nature of their selecting ligand in the thymus is not known. However, it is proposed that the lysosomal enzyme α-galactosidase A could promote generating the endogenous lipid antigens necessary for iNKT cell selection and activation (5). iNKT cell generation also requires homotypic interaction of the costimulatory molecules, Slamf1 and Slamf6, which are highly expressed on DP cells (6). Thus, it is currently understood that immature iNKT cells are selected by DP thymocytes which present the selecting ligand and co-stimulatory signals for positive selection. Consequently, iNKT cells substantially differ in their thymic developmental pathway from conventional αβ T cells which require thymic epithelial cells for their selection.
After their positive selection, thymic iNKT cells undergo a series of phenotypic changes that are associated with acquisition of distinct effector function (78). Depending on their signature cytokine production and subset-specific transcription factor expression, 3 distinct populations of iNKT cells have been identified, and they are referred to as NKT1, NKT2, and NKT17 cells (9). The developmental mechanism and functional aspects of these iNKT subsets, however, remain largely unknown, and they represent a hotly contended area of research (10). The common γ-chain family cytokine, IL-15, is necessary for the terminal maturation and the generation of T-bet-expressing NKT1 cells (1112). The cellular signals that are required for development of other iNKT subsets, however, remain mostly unknown. Syndecan-1 was recently proposed to be specifically required for NKT17 cell generation (13), but it is not known why. Along these lines, distinct expression of the highly glycosylated isoform of CD43 and ICOS (CD278) were found to be associated with iNKT subset differentiation (14), but again the molecular basis of their distinct expression also remains obscure.
While mapping the signaling requirements of iNKT subset differentiation is of great interest, the scarcity of these cells in the thymus (<1.0% among all thymocytes) makes it difficult for in-depth analysis (115). Pre-enrichment of iNKT cells to obtain increased numbers of cells for analysis can greatly facilitate their further downstream examination, such as for cell sorting, flow cytometric analysis or various in vitro manipulations (1617181920). Because iNKT cells specifically bind to glycolipid-antigen loaded CD1d tetramer molecules (CD1dTet) (212223), enrichment of CD1dTet+ cells is frequently used to facilitate the analysis iNKT cells (1617181920). Indeed, we confirmed that magnetic-activated cell sorting (MACS)-assisted isolation of CD1dTet+ cells dramatically enriched of iNKT cells from BALB/c thymocytes, corroborating the effectiveness of this method. Surprisingly, however, we also found that MACS-based enrichment of iNKT cells introduced a severe bias into the subset composition of recovered iNKT cells. Thus, here, we reassessed the efficacy of the MACS-based iNKT enrichment method, and we report a previously unappreciated effect of CD1dTet-mediated selection of iNKT cells that preferentially enriches for NKT2 lineage cells and results in loss of NKT1 cells.
MATERIALS AND METHODS
Mice
BALB/c and C57BL/6 (B6) mice of both sexes were obtained from the Charles River Laboratories (Frederick, MD, USA), and analyzed between 6–12 wk of age. Animal experiments were approved by the National Cancer Institute (NCI) Animal Care and Use Committee and all animal experiments were performed as approved. All mice were cared for in accordance with National Institutes of Health (NIH) guidelines.
MACS-based enrichment of iNKT cells
Total thymocytes were processed to single cell suspension in 0.5% BSA, 2 mM EDTA in PBS (20×106 cells/ml) and incubated with phycoerythrin (PE)-conjugated CD1d-Tetramers for 60 min on ice. Unbound reagents were washed out with 0.1% BSA, 2 mM EDTA in PBS, and cell pellets were resuspended in the same buffer to 100×106 cells/ml. Anti-PE MACS beads (Miltenyi Biotec, Bergisch Gladbach, Germany) were then added to the cell suspension at a ratio of 200 µl beads per 100×106 cells, and incubated for 20 min at 4°C. Excess antibodies were removed by washing cells with 0.5% BSA, 2 mM EDTA in PBS and then resuspended in the same buffer to a concentration of 200×106 cells/ml. Positively labeled cells were selected by magnetic separation using LS columns, following the manufacturer's instruction (Miltenyi Biotec).
Magnetic-activated depletion of CD24+ thymocytes
Total thymocytes and lymph node cells were processed to single cell suspension in 10% FBS/PBS (20×106 cells/ml), and incubated with rat anti-mouse CD24 antibodies (2.5 μg/5×106 cells) for 30 min on ice. Excess antibodies were washed out with 10% FBS/PBS. Each 100×106 cells were mixed with 3 ml of anti-rat IgG conjugated BioMag beads (QIAgen, Frederick, MD, USA). After one hour of incubation at 4°C on a MACSmix Tube Rotator (Miltenyi Biotec), anti-CD24 antibody-bound cells were magnetically depleted, and non-binding cells were used for further experiments.
Flow cytometry
Flow cytometry data were acquired on LSRFortessa or LSR II flow cytometers (BD Biosciences, San Jose, CA, USA) and analyzed using software designed by the Division of Computer Research and Technology, NCI. Live cells were gated using forward scatter exclusion of dead cells stained with propidium iodide. Fixation and permeabilization were performed with Foxp3 Transcription Factor Staining Buffer kit according to the manufacturer's instructions (Thermo Fisher eBioscience, Waltham, MA, USA). Antibodies with the following specificities were used for staining: TCRβ (H57-597), CD24 (M1/69), and T-bet (eBio4B10), were from Thermo Fisher eBioscience; retinoid-related orphan receptor gamma t (RORγt; Q31-378), B220 (RA3-6B2), CD69 (H1.2F3), CD5 (53-7.3), were from BD Biosciences; promyelocytic leukemia zinc finger (PLZF; 9E12) was from BioLegend (San Diego, CA, USA). CD1d tetramers loaded with PBS-57 and unloaded controls were obtained from the NIH tetramer facility (Emory University, Atlanta, GA, USA) and used to identify thymic iNKT cells as previously described (24).
In vitro stimulation of iNKT cells
iNKT cells from total or CD24+ cell-depleted B6 splenocytes were stimulated in vitro using PMA (25 ng/ml) and ionomycin (1 µM, both from Sigma) for 4 hours. After stimulation, excess reagents were washed out with cell staining buffer (0.1% BSA, 0.1% sodium azide in HBSS), and cells were stained for surface marker expression.
RESULTS AND DISCUSSION
MACS-based selection of iNKT cells results in preferential enrichment for NKT2 cells
Glycolipid-loaded CD1d tetramers represent an effective tool to identify iNKT cells that express the canonical Vα14-Jα18 TCR (1521). In the thymus of BALB/c mice, usually less than 1% of total thymocytes correspond to CD1dTet+ iNKT cells (Fig. 1A top) (18). Notably, not all of them are functionally mature, and a significant proportion of these cells correspond to immature pre-selection CD24hi iNKT cells (Fig. 1A, bottom). Because of their scarcity, enrichment of thymic iNKT cells greatly facilitates their subsequent analysis. To this end, MACS-based positive selection of CD1d+ iNKT cells from total thymocytes is frequently employed to increase frequencies of iNKT cells and to facilitate detailed interrogation of their phenotype and function (1617181920). We confirmed that this protocol indeed dramatically enriched for iNKT cells (Fig. 1B), usually resulting in an approximately 90±18.7-fold increase in iNKT cell frequencies (Fig. 1C). The flow-through fraction of MACS columns, on the other hand, showed substantially decreased frequencies of iNKT cells, indicating preferential binding of MACS-bead labeled iNKT cells to magnetized MACS columns (Supplementary Fig. 1A). Interestingly, we also noticed a dramatic shift in TCRβ surface expression and in the amount of CD1dTet+ binding by post-enrichment iNKT cells (Fig. 1D). Compared to pre-enrichment iNKT cells, MACS-selected iNKT cells expressed greater amounts of TCRβ and showed increased staining for CD1dTet reagents (Fig. 1D). These results suggested that CD1dTet-mediated retention of iNKT cells in MACS columns has the unintended effect of enriching for iNKT cells with larger amount of surface TCRβ expression and greater CD1dTet-binding capacity. Along these lines, we found that the unselected flow-through fraction still contained few iNKT cells, but that they expressed much smaller amounts of TCRβ and showed decreased binding of CD1dTet (Supplementary Fig. 1B). Thus, CD1dTet-binding MACS columns act as a cellular sieve which preferentially enriches for iNKT cells that bind greater amounts of CD1dTet. Collectively, these results indicated that MACS-based selection of CD1dTet+ cells introduces a bias during the enrichment of iNKT cells, so that iNKT cells expressing higher levels of surface TCRβ are preferentially retained.
Figure 1
CD1d-tetramer-based enrichment of thymic iNKT cells. (A) Identification of iNKT cells in BALB/c thymocytes by CD1d tetramer (CD1dTet) vs. TCRβ (top) or CD1dTet vs. CD24 analysis (bottom). Results are representative of 5 independent experiments. (B) MACS-based enrichment of CD1dTet+ iNKT cells is demonstrated by CD1dTet vs. TCRβ (top) or CD1dTet vs. CD24 analysis (bottom) of iNKT cells in total thymocytes or after MACS column enrichment. Results are representative of 5 independent experiments. (C) Percentages of iNKT cells in total thymocytes (before) and CD1dTet-enriched fraction (after). Plot shows summary of 5 independent experiments. (D) Surface TCRβ expression and CD1dTet staining on thymic iNKT cells before and after MACS-mediated enrichment for iNKT cells. Histograms (left) are representative and graphs (right) show summary of 5 independent experiments. (E) Intranuclear staining for PLZF and RORγt shows subset distribution before and after MACS-mediated enrichment for thymic iNKT cells. Enriched iNKT cells were stained for CD24 and gated on CD24lo to identify mature iNKT cells. Dot plots (left) are representative and graphs (right) show summary of 5 independent experiments.
NS, not significant.
**p<0.01; ***p<0.001 were considered statistically significant.
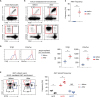
The amount of surface TCRβ and binding of CD1dTet differ among individual iNKT subsets (25). Thus, we wished to examine if MACS-based iNKT enrichment would also skew the subset composition of enriched iNKT cells, when compared to that of pre-enrichment iNKT cells. Individual iNKT subsets can be identified by the distinct expression of 3 transcription factors, namely PLZF, RORγt, and T-bet (926). NKT1 cells express low amounts of PLZF but high levels of T-bet. NKT2 cells, on the other hand, are abundant for PLZF but not for RORγt or T-bet. Finally, NKT17 cells express the signature transcription factor RORγt, and they are absent for T-bet (927). Here, we found that MACS-enrichment for CD1dTet+ cells induced a significant bias for NKT2 lineage cells, with a concomitant loss in NKT1 cells (Fig. 1E). The unbound fraction of iNKT cells that were recovered from the flow-through of the MACS column, on the other hand, showed a reverse enrichment for TCRβlo NKT1 lineage cells and selective loss of TCRβhi NKT2 cells (Supplementary Fig. 1C). Collectively, these results unveil a previously unappreciated pitfall in MACS-based iNKT enrichment protocols that skews the subset composition of recovered iNKT cells into NKT2 lineage cells. Because of this limitation, we found it necessary to develop an alternative approach to enrich iNKT cells, and to develop a method that would not employ anti-TCRβ or CD1dTet for enrichment.
Magnetic-activated depletion of CD24+ thymocytes permits effective iNKT cell enrichment
CD24 is a glycosylphosphatidylinositol (GPI)-linked surface protein that is highly expressed on immature thymocytes (282930). Positive selection in the thymus and further maturation of thymocytes result in loss of CD24 expression, so that all mature αβ T cells, including iNKT cells, have downregulated CD24 surface expression (1). By removing CD24+ cells from total thymocytes, we expected that we would enrich for iNKT cells, and specifically enrich for mature iNKT cells that do not express CD24. This was indeed the case (Fig. 2A). Using anti-CD24 antibodies followed by magnetic-activated depletion by BioMag beads, we found that this procedure routinely resulted in a 22±3.71-fold enrichment of iNKT cells from total thymocytes of BALB/c mice (Fig. 2B, left). We obtained similar results when enriching for iNKT cells from C57BL/6 thymocytes (Supplementary Fig. 2A). Notabley, the overall recovery rate of enriched iNKT cells was 19%±5.19% of the expected number of iNKT cells from total BALB/c thymocytes. These numbers were statistically not different from those when using the CD1dTet-based MACS enrichment method (Fig. 2B, right). However, and in marked contrast to the results from CD1dTet-based MACS enrichment (Fig. 1E), we did not find any significant changes in CD1dTet staining or surface TCRβ expression after their enrichment (Fig. 2C). Because the surface phenotype for these 2 markers was preserved after CD24+ thymocyte depletion, these results suggested that CD24-depletion could represent a superior alternative to iNKT cell enrichment by CD1dTet-binding and MACS.
Figure 2
Enrichment of mature thymic iNKT cells by depletion of CD24+ thymocytes. (A) Enrichment of thymic iNKT cells by magnetic activated depletion of CD24+ thymocytes. Dot plots (left) are representative and graph (right) shows summary of 5 independent experiments. (B) Comparison of fold-enrichment and iNKT recovery rates of MACS vs. CD24+ thymocytes depletion methods to enrich thymic iNKT cells. Fold enrichment was determined by dividing the frequency of iNKT cells before enrichment by the frequency of iNKT cells after MACS enrichment. Recovery rate was defined as the percentage of iNKT cell number that was recovered after enrichment compared to the total estimated iNKT number in the starting population. Results are summary of 4 independent experiments. (C) Surface TCRβ expression and CD1dTet staining on thymic iNKT cells before and after magnetic activated depletion of CD24+ thymocytes. Histograms are representatives of 5 independent experiments. (D) iNKT subset composition in unfractionated BALB/c thymocytes (before), and CD24+ cell-depleted thymocytes (after). Dot plots (left) are representative and graph (right) shows summary of 6 independent experiments.
NS, not significant.
*p<0.05; ***p<0.001 were considered statistically significant.

To examine if this alternative enrichment method would indeed permit maintenance of the iNKT subset composition, next, we performed intranuclear staining of iNKT cells before and after CD24+ cell depletion. Notably, the overall composition of iNKT subsets remained mostly preserved (Fig. 2D), and there was no selective loss of NKT2 lineage cells, as previously observed for CD1dTet-based MACS enrichment (Fig. 1E). We also obtained similar results when enriching iNKT cells from C57BL/6 thymocytes, indicating that CD24+ cell depletion can effectively enrich iNKT cells independently of the mouse strain (Supplementary Fig. 2B). An additional advantage of this new method is the removal of immature iNKT cells which are CD24+ (Stage 0), so that it results in selective enrichment of CD24-negative mature iNKT cells. Also, because some iNKT cells do express surface CD8 coreceptors (31), enriching iNKT cells by CD24+ cell depletion is advantageous over depletion of CD8+ thymocytes (32), because CD8+ cell depletion could inadvertently remove such CD8+ iNKT cells from the pool. Notably, CD24+ depletion also permitted enrichment for iNKT cells in peripheral tissues, because it removed B cells from the cell suspension. Unlike mature T cells, peripheral B cells express large amounts of CD24 (Fig. 3A) (33). Thus, CD24+ B cell depletion significantly enriched iNKT cells from spleen cells (Fig. 3B), and such iNKT cells were functionally intact and responded to activating signals comparable to spleen iNKT cells prior to enrichment (Fig. 3C).
Figure 3
Enrichment of mature peripheral iNKT cells by depletion of CD24+ splenocytes. (A) Identification of CD24+ cells by surface TCRβ and B220 staining. Dot plots are representative of 2 independent experiments. (B) Mature iNKT cell enrichment from C57BL/6 spleen cells by magnetic-activated depletion of CD24+ cells. Dot plots (top) are representative and graph (bottom) shows summary of 3 independent experiments. (C) Surface CD69 and CD5 staining on total and CD24+ cell depleted BALB/c spleen iNKT cells upon PMA + ionomycin stimulation. Results are representative of 2 independent experiments.
**p<0.01 were considered statistically significant.

Finally, another major benefit of using this procedure is the ability to procure iNKT cells that have not been exposed to CD1d tetramers. CD24-depleted iNKT cells remain untouched in their TCR, because they were not selected based on their TCR binding to CD1d tetramers. Consequently, this method avoids unintended activation of iNKT cells by CD1dTet engagement during their enrichment process, and can be used to enrich thymic iNKT cells before proceeding to electronic sorting (Supplementary Fig. 3), which dramatically shortens the sorting time and increases viability of the sorted iNKT cells.
In summary, here we report a previously unappreciated drawback in isolating iNKT cells using glycolipid-loaded CD1d tetramers. We found that this conventional procedure inadvertently enriches for iNKT cells expressing large amounts of TCRβ and binding greater quantities of CD1d-tetramers (TCRβhi CD1dTethi), which mostly correspond to NKT2 lineage cells (27). Consequently, assessing the cytokine expression profile or other effector functions of such enriched iNKT cells need to consider the shortcomings of this method, and interpret the results in this context. Because iNKT cells are scarce in the thymus (1), securing an effective method to enrich iNKT cells from a given cell population is pivotal for their correct and detailed analysis. We propose magnetic-activated depletion of CD24+ thymocytes as a reliable method to achieve this goal, and we validated its efficacy by analyzing iNKT subsets in thymocytes before and after CD24+ cell depletion.
 XML Download
XML Download