

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Hematopoietic stem cells (HSCs) are pluripotent cells that reside in the bone marrow and can differentiate into all blood cell lineages. The maintenance of HSCs is essential to ensure hematopoiesis is viable for the life of the organism. The HSC niche, a specific microenvironment where these cells form and reside, plays a pivotal role in maintaining HSCs via cell–cell contact and/or production of chemokines and cytokines (12). Furthermore, cell-intrinsic factors, such as GATA-2 (3) and Bim-1 (45) regulate HSC self-renewal and quiescence, and the anti-apoptotic Bcl-2 family proteins are required for survival of HSCs under stress conditions (67). Recently, several studies revealed that autophagy is related to the maintenance of HSCs attributing to its pivotal role in cellular homeostasis and cell survival. Autophagy is a highly conserved lysosome-dependent degradation pathway in eukaryotes and is used to maintain homeostasis by degrading old or damaged cellular proteins and organelles (8910). mTOR complex 1, which is negative regulator of autophagy, is essential for regulation of HSC quiescence (1112), and autophagy-regulating transcription factor Forkhead box O3a induces autophagy to protects HSCs from metabolic stress (13). Previously, it has been suggested that the essential autophagy machinery component Atg7 is required for the maintenance of HSC integrity, production of both lymphoid and myeloid progenitors, and for suppression of myeloproliferation (14). Also, hematopoietic cell specific Atg7 deficiency lead to altered erythroid developmental stages and lethal anemia (15). Furthermore, another essential autophagy molecule Atg5 plays an important role in B cell development (16), plasma cell differentiation (17), development of innate lymphocytes (18) and is associated with acute myeloid leukemia (19); however, the role of Atg5 in the self-renewal and differentiation of HSCs has not been investigated thoroughly.
Here, we demonstrated the role of Atg5 as a regulator for maintaining the number and proliferation of HSCs. Atg5 deficiency in HSCs resulted in a survival defect with severe lymphopenia and anemia. The absence of Atg5 results in aberrant proliferation of Lin−Sca-1+c-Kit+ (LSKs) and significant reduction of HSCs, mature progenitors, and terminally differentiated cells. Furthermore, the reconstitution ability of HSCs was significantly decreased following hematopoietic cell-specific Atg5 deficiency. Our findings suggest that Atg5 plays a crucial role in the maintenance and reconstitution ability of HSCs.
MATERIALS AND METHODS
Mice
Mice were housed in a specific pathogen-free facility at Korea Advanced Institute of Science and Technology (KAIST). Vav-iCre mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA), and Atg5flox/flox mice (20) were gifted from Akiko Iwasaki (Yale University, New Haven, CT, USA). Vav_Atg5-/- mice were obtained by crossing the Vav-iCre mice and Atg5flox/flox mice, and their genotypes were confirmed using tail genomic DNA. Littermate Atg5flox/flox mice were used as control mice. In all experiments, sex- and age-matched mice between 7–14 weeks of age were used. All animal procedures were approved by and performed according to the standards of the Institutional Animal Care and Use Committee of KAIST (KA2016-18).
Bone marrow, spleen, lymph node, and blood isolation
Mice were euthanized with carbon dioxide gas, and bone marrow cells were isolated from tibias and femurs of hind legs from WT and Vav_Atg5−/− mice using a syringe with DMEM containing 1% FBS (Welgene, Daegu, Korea). The lymph nodes were removed, minced using a razor, and incubated in PBS containing 1% FBS with 2 mg/ml of collagenase IV (Worthington Biochemical Corporation, Lakewood, NJ, USA) and 30 μg/ml of DNase I (Roche, Basel, Switzerland) for 30 min at 37°C. Then cells were centrifuged at 1,500 rpm for 5 min at 4ºC and treated with HBSS buffer containing 5% FBS and 5 mM EDTA for 5 min at 37°C. Lymph nodes and spleens were disrupted through a 70-μm cell strainer and plunger using a syringe. After isolation, all cells were treated with ammonium-chloride-potassium lysis buffer for 5 min at room temperature to remove the RBCs. Whole blood cell samples were obtained from hearts using a syringe in 4% trisodium citrate buffer to prevent blood solidification. The isolated cells were counted using a hematocytometer following staining with 1% trypan blue (SPL Life Sciences, Pocheon, Korea) for exclusion of the dead cells.
Flow cytometry
For flow cytometry analysis, cells were stained with the indicated antibodies. FITC anti-mouse CD3 (17A2; BD Bioscience, Franklin Lakes, NJ, USA), CD11b (M1/70; BD Bioscience), B220 (RA3-6B2; BD Bioscience), Ly6C (AL-21; BD Bioscience), and Ly6G (1A8; BD Bioscience) antibodies were used as lineage markers. PE anti-mouse CD48 (HM48-1; BD Bioscience), PerCPCy5.5 anti-mouse Sca-1 (D7; BioLegend, San Diego, CA, USA), PECy7 anti-mouse c-Kit (CD117, 2B8; eBioscience, San Diego, CA, USA), APC anti-mouse CD150 (TC15-12F12.2; BioLegend), biotin anti-mouse Flt3 (A2F10; BioLegend), APC anti-mouse CD127 (IL-7Rα, A7R34; BioLegend), Alexa Fluor 647 anti-mouse CCR9 (CW-1.2; BioLegend), PE anti-mouse CD122 (TM-β1; BioLegend), PECy7 anti-mouse NK1.1 (PK136; eBioscience), APC anti-mouse DX5 (CD49b, DX5; BD Bioscience), PE anti-mouse CD105 (MJ7/18; BioLegend), PECy7 anti-mouse CD41 (MWReg30; BioLegend), APCCy7 anti-mouse CD16/CD32 (93; BioLegend), biotin anti-mouse c-Kit (2B8; BD Bioscience), streptavidin-PE (BD Bioscience), and streptavidin-Alexa Fluor 700 (Life Technologies, Carlsbad, CA, USA) antibodies as well as DAPI were used to characterize the hematopoietic progenitor cells. PerCPCy5.5 anti-mouse F4/80 (BM8; eBioscience), APCCy7 anti-mouse CD11b (M1/70; BD Bioscience), Alexa Fluor 647 anti-mouse MHC class II (M5/114.15.2; BD Bioscience), APC anti-mouse CD11b (M1/70; BD bioscience), PE anti-mouse Siglec-F (E50-2440; BD bioscience), PerCPCy5.5 anti-mouse Ly6G (1A8; BD bioscience), PECy7 anti-mouse CD11c (N418; BioLegend), APCCy7 anti-mouse Ly6C (AL-21; BD Bioscience), FITC anti-mouse CD3, PE anti-mouse CD8a (53-6.7; BioLegend), PerCPCy5.5 anti-mouse B220 (RA3-6B2; BD Bioscience), and APC anti-mouse CD4 (GK1.5; BioLegend) antibodies as well as DAPI were used to analyze immune cell populations according to previously described methods (21).
For intracellular staining, cells were stained with biotin anti-mouse lineage markers (CD3e, CD11b, Gr-1, B220 (BD Bioscience), FITC or PerCPCy5.5 anti-mouse Sca-1 (BioLegend), PECy7 anti-mouse c-Kit, and streptavidin-APCCy7 (BD Bioscience) antibodies. After washing by FACS buffer, the cells were stained with FITC labeled Annexin V (BD Bioscience), or cells were fixed and permeabilized with Fixation/Permeabilization Solution Kit (BD Biosciences) and stained with PE anti-mouse 53BP1 (Novus Biologicals, Centennial, CO, USA) or Ki-67 (eBioscience) antibodies. For mitochondrial staining, MitoTracker Green, MitoTracker Red, and MitoSox (Thermo Fisher, Waltham, MA, USA) were used according to manufacturer's instructions. Flow cytometry experiments were performed on a FACS Fortessa (BD Biosciences), and data were analyzed using FlowJo software (Tree Star, Inc., San Diego, CA, USA).
BrdU incorporation assay
Mice were injected intraperitoneally with 1 mg/ml of BrdU (BD Biosciences) in 200 μl DPBS. After 24 h, the mice were sacrificed, and bone marrow cells were isolated. BrdU staining was performed according to the manufacturer's instructions. For surface staining, 2×106 bone marrow cells were stained with PE anti-mouse CD11b, CD3, B220, Ly6C, and Ly6G antibodies (BD bioscience), PerCPCy5.5 anti-mouse Sca-1 antibodies, and PECy7 anti-mouse c-Kit antibodies for 30 min on ice. Cells were examined using FACS Fortessa, and data were analyzed with FlowJo software (Tree Star, Inc.).
Bone marrow reconstitution
A total of 2×106 bone marrow cells were transplanted intravenously into lethally irradiated CD45.1+ mice. Eight wk after transplantation, reconstitution of recipient bone marrow cells was confirmed in peripheral blood using PE anti-mouse CD45.1 (A20; BD Bioscience) and APC anti-mouse CD45.2 (104; BD Bioscience) antibodies.
RESULTS
Atg5 deficiency in hematopoietic cells causes survival defects in mice
Because Atg5 deficiency in mice is neonatally lethal, we generated hematopoietic cell-specific Atg5 knockout mice to elucidate the role of Atg5 during hematopoiesis. As Vav, one of the guanine nucleotide exchange factors, is expressed in hematopoietic cells regardless of lineage or developmental stage, we crossed Atg5flox/flox mice with Vav-iCre animals to generate Vav_Atg5−/− mice and monitored the survival of newborn pups. Vav_Atg5−/− mice died within 11 wk (Fig. 1A) and exhibited body weights that were lower than control mice (Fig. 1B). These results indicated that deficiency in Atg5 in hematopoietic systems leads to a survival defect. We hypothesized that the decreased survival of Vav_Atg5−/− mice is attributable to the impairment of hematopoiesis, because hematopoietic cell-specific Atg7 deficiency developed severe anemia and lymphopenia (14). To determine the effect of Atg5 on hematopoiesis, we assessed total cell numbers in bone marrow, spleen, and lymph nodes. As predicted, Vav_Atg5−/− mice displayed significantly diminished total cell numbers in these compartments compared to control mice (Fig. 1C).
 | Figure 1Vav_Atg5−/− mice display defects in survival, body weights, and total hematopoietic cell number. Starting 3 wk after birth, survival (A) and body weights (B) of newborn control and Vav_Atg5−/− pups were monitored every wk. (C) The number of bone marrow, spleen, and lymph node cells from these animals were counted (n=3). Data are representative of 3 independent experiments and presented as mean ± SEM.
*p<0.05, **p<0.01, ***p<0.001 as calculated by Student's t-test.
|
Hematopoietic cell-specific Atg5-deficient mice suffer from severe lymphopenia
Next, we analyzed the immune cell populations in Vav_Atg5−/− mice. T cells were significantly reduced in spleens (Fig. 2A and B) and lymph nodes (Fig. 2C and D), while B cells were diminished only in spleens (Fig. 2A and B). Myeloid cells were slightly increased in spleens (Fig. 2A and B) and lymph nodes (Fig. 2C and D) compared to control mice, but these differences failed to reach statistical significance except for the neutrophils in lymph nodes (Fig. 2C and D). These results indicated that Atg5 is crucial for the development of lymphoid cells and that hematopoietic cell-specific Atg5 gene deletion causes severe lymphopenia.
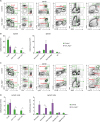 | Figure 2Vav_Atg5−/− mice suffer from severe lymphopenia. Spleen (A, B) and lymph nodes (C, D) were isolated from control and Vav_Atg5−/− mice. B cells (CD3−B220+), CD4 (CD3+CD4+) T cells, CD8 (CD3+CD8a+) T cells, dendritic cells (DC; B220−CD11C+MHC class II+) in spleen, resident DCs (B220−CD11c+MHC class IIInt) and migratory DCs (B220−CD11c+MHC class IIHigh) in lymph nodes, neutrophils (CD11b+Ly6G+), monocytes (CD11b+Ly6CHigh), and macrophage (CD11b+F4/80+) were classified by flow cytometry. Representative FACS plots (A, C) and absolute cell numbers (B, D) were calculated (n=3). Data are representative of three independent experiments and presented as mean ± SEM.
*p<0.05 as calculated by Student's t-test.
|
Atg5 deficiency results in defective erythropoiesis
Next, we examined erythropoiesis in Vav_Atg5−/− mice, because the overall hematopoiesis includes lymphopoiesis, myelopoiesis, and erythropoiesis. Previous study has reported that Atg5 was highly expressed in the pro-erythroblast and the expression levels were gradually reduced depending on the stage of erythropoiesis (15), suggesting that Atg5 is potentially associated with erythroid maturation. RBCs in peripheral blood were counted using a hemocytometer, and bone marrow and spleen samples were stained with CD71 and Ter119 to assess the developmental stage of the erythrocytes. Vav_Atg5−/− mice showed a reduced number of RBCs in peripheral blood compared with control animals (Fig. 3A), indicating that Vav_Atg5−/− mice suffer from anemia. The distribution of erythrocyte developmental stages was also altered in Vav_Atg5−/− mice compared with controls (Fig. 3B). Specifically, basophilic erythroblasts were decreased concomitant with an increase in the proportions of other stages (Fig. 3C). These results indicate that Atg5 deficiency in hematopoietic cells leads to altered erythroid differentiation and causes RBC reduction.
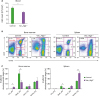 | Figure 3Erythropoiesis is altered following Atg5 deficiency. (A) The number of RBCs in peripheral blood were counted in control and Vav_Atg5−/− mice. (B) Erythroid developmental stages in bone marrow and spleen were assessed by flow cytometry. Pro-erythroblasts (Pro_ery; Ter119−CD71High), basophilic erythroblasts (Baso_ery; Ter119+CD71High), chromatophilic erythroblasts (Chro_ery; Ter119+CD71Med), and orthochromatophilic erythroblasts (Ortho_ery; Ter119+CD71−) were characterized (n=4). Data are representative of three independent experiments and presented as mean ± SEM.
*p<0.05 and **p<0.01 as calculated by Student's t-test.
|
Atg5 is essential for the HSCs maintenance and reconstitution ability
In bone marrow, hematopoietic cells are divided into Lin−c-Kit+ (LK) cells enriched for myeloid progenitors and megakaryocyte-erythroid progenitors and LSK cells comprising long-term HSCs, short-term HSCs, and multipotent progenitors (2223). Therefore, we analyzed LK and LSK populations to clarify the developmental impairment of HSCs and hematopoietic progenitor cells in Vav_Atg5−/− mice. LSKs in bone marrow were increased in Vav_Atg5−/− mice, while LKs were significantly decreased in these mice compared with controls (Fig. 4A). We then analyzed HSCs and other hematopoietic progenitors in bone marrow. HSCs were characterized as LSK CD48−CD150+ cells. Interestingly, HSCs in bone marrow of Vav_Atg5−/− mice were significantly reduced compared with controls, indicating that reduced HSC numbers were not attributable to the LSK expansion (Fig. 4B).
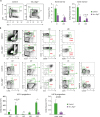 | Figure 4Atg5 deficiency leads to defective maintenance of HSCs and developmental impairment of hematopoietic progenitor cells. (A, B) Bone marrow cells isolated from 11-wk-old from control and Vav_Atg5−/− mice were analyzed. (A) Frequency and absolute number of LK and LSK cells were assessed by flow cytometry (n=4–5). (B) HSCs were characterized as LSK CD48−CD150+, and multiple progenitor cells were characterized in CLPs (Lin−IL-7Ra+Flt3+), CCR9+ LMPPs (LSK Flt3+), NKPs (Lin−CD122+NK1.1−DX5−), MkPs (LK CD41+CD150+), and GMPs (LK CD41−FcgRII/III+) (n=3). Data are representative of three independent experiments and presented as mean ± SEM.
*p<0.05, **p<0.01, ***p<0.001 as calculated by Student's t-test.
|
For the lymphoid progenitor compartment, lymphoid-primed multipotent progenitors (LMPPs), common lymphoid progenitors (CLPs), and NK cell progenitors (NKPs) were analyzed, while megakaryocyte progenitors (MkPs) and granulocyte-macrophage progenitors (GMPs) were analyzed for the myeloid progenitor compartment. In Vav_Atg5−/− mice, NKPs, and GMP were significantly reduced (Fig. 4B), while CLPs, CCR9+ LMPPs, and MkPs were diminished but without statistical significance. These results indicate that Atg5 is crucial for the maintenance of HSCs and that a defect in HSCs results in developmental impairment of hematopoietic progenitor cells in Vav_Atg5 mice.
Next, we assessed proliferation and apoptosis in Atg5-deficient hematopoietic cells. Bone marrow cells were stained with anti-Ki67, BrdU antibodies and Annexin V. As shown in Fig. 5A, the LSK compartment in Vav_Atg5−/− mice exhibited enhanced resistance to apoptosis and higher proliferation rates compared with controls. Specifically, the Annexin V+ populations in LSK cells were reduced in Vav_Atg5−/− mice. As Annexin V binds phosphatidylserine on the apoptotic cell surface, the reduction of Annexin V+ populations suggests that LSK apoptosis was decreased in Vav_Atg5−/− mice. Furthermore, BrdU+ populations and Ki67 levels were enhanced in LSKs of Vav_Atg5−/− mice, suggesting that hematopoietic cell-specific Atg5 deficiency results in aberrant proliferation of LSKs.
 | Figure 5Hematopoietic cell specific Atg5 deficiency results in aberrant proliferation and mitochondrial dysfunction in hematopoietic cells. (A) Apoptosis and cell proliferation assays of LSKs from control and Vav_Atg5−/− mice were performed using Annexin V, BrdU, and Ki67 staining. For the BrdU assay, proliferation of LSK cells was analyzed 24 h after BrdU injection (n=4–5). (B) Approximately 2×106 of total bone marrow cells from control and Vav_Atg5−/− mice were injected intravenously into lethally irradiated CD45.1+ mice. After 8 weeks, percentage of reconstitution of transplanted bone marrow in peripheral blood was confirmed by flow cytometry (n=3). (C) Mitochondrial functions in bone marrow LSK cells from control and Vav_Atg5−/− mice were assessed using MitoTracker Green, MitoTracker Red, and MitoSox (n=4–5). Data are representative of 3 independent experiments and presented as mean ± SEM.
*p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 as calculated by Student's t-test.
|
In addition, we assessed the reconstitution ability of HSCs with Atg5 deficiency. Approximately 2×106 bone marrow cells from control and Vav_Atg5−/− (CD45.2+) mice were injected via tail vein into lethally irradiated CD45.1+ congenic mice. After 8 wk, more than 90% of cells were reconstituted in peripheral blood from the recipients transplanted with control bone marrow cells, while the Vav_Atg5−/− bone marrow cells led to reconstitution of about 70% of the cells in the hematopoietic system of recipients (Fig. 5B). These results indicate that Atg5 affects not only the maintenance but also the reconstitution ability of HSCs.
Autophagy contributes to the maintenance of intracellular homeostasis via removal of damaged mitochondria, and the absence of autophagy leads to accumulation of dysfunctional mitochondria and ROS. Therefore, we hypothesized that Atg5 deficiency in hematopoietic cells results in mitochondrial disturbance. We analyzed LSK populations in bone marrow to determine whether Atg5 deficiency mediates mitochondrial dysfunction in hematopoietic cells. We assessed the mitochondrial functions in bone marrow LSK cells using MitoTracker Green, MitoTracker Red, and MitoSox. As expected, LSK cells showed enhanced mitochondrial mass and membrane potentials in Vav_Atg5−/− mice, while mitochondrial superoxide levels were slightly, but not significantly, increased (Fig. 5C). These results indicate that Atg5 deficiency causes unregulated accumulation of mitochondria in LSKs.
DISCUSSION
Autophagy is crucial for the maintenance of cellular homeostasis via elimination of impaired cytosolic components, such as damaged mitochondria and misfolded proteins, and this process therefore plays an essential role in cell survival (24). During the early neonatal starvation period, autophagy promotes survival of newborn pups by degrading self-proteins to supply amino acids (25). In mesenchymal stromal/stem cells, autophagy is induced under hypoxic condition and promotes proangiogenic activity via enhancing angiogenin and VEGF production (26). Furthermore, autophagy and autophagy-related proteins have been reported as crucial regulators for hematopoiesis. For example, Atg7 is known to play an important role in the maintenance of HSCs, and Atg7 deficiency in hematopoietic cells causes severe anemia and lymphopenia in vivo (1415). Also, Atg5 is required for the development and survival of innate lymphoid cells and NK cells, as this factor facilitates lymphocyte survival following homeostasis proliferation during lymphopenia (18); however, the role of the autophagy protein Atg5 in HSCs has not previously been thoroughly investigated.
In this study, we found that hematopoietic cell-specific Atg5 deficiency causes severe lymphopenia, anemia, and survival defects. Immune cell populations in spleens and lymph nodes were significantly altered in Vav_Atg5−/− mice, due to the developmental impairment of hematopoietic progenitor cells. Specifically, Vav_Atg5−/− mice showed defective HSC maintenance and lower reconstitution capacity following disrupted differentiation of hematopoietic progenitor cells. Also, abnormal distribution of erythrocyte developmental stages resulted in decreased RBCs in blood in Vav_Atg5−/− mice, and hematopoietic cell-specific Atg5 deficiency led to aberrant proliferation of LSKs. Although LSK cells displayed enhanced proliferation and diminished apoptosis, absolute number of HSCs were significantly decreased in Vav_Atg5−/− mice. We propose that defective reconstitution ability of bone marrow cells from Vav_Atg5−/− mice are attributed to the lower HSC numbers. Also, autophagy is required for the DNA damage responses and loss of autophagy facilitate the accumulation of damaged DNA inducing genetic instability (27282930), therefore, enlarged LSK cells do not necessarily reflect the abundance of functional HSCs. Finally, we found that Atg5-deficient LSKs displayed unregulated accumulation of mitochondria. Mitochondria mass and membrane potentials were enhanced in LSKs of Vav_Atg5-/- mice. Because autophagy eliminates damaged organelles, such as mitochondria, these findings suggest that loss of Atg5 causes impairment of autophagy function and ultimately enhances the intracellular stress in hematopoietic cells. Mitochondria, which is the main producer of ROS in cells (31), continuously repeat fusion and fission during the life cycle to control mitochondrial quality (32), and mitochondrial fusion helps to redistribute metabolites and proteins and to dilute damage materials to reduce mitochondrial stress (33). Despite disruption of mitochondrial membrane potential results in release of cytochrome C into the cytosol and apoptosis, mitochondrial hyperpolarization also lead to cellular abnormalities. For example, patients with systemic lupus erythematosus exhibited mitochondrial hyperpolarization (3435). Therefore, we suggest that increased mitochondrial mass and membrane potential indicates enhanced mitochondrial stress accumulation that inhibits cellular function and survival. Further, although the differences were not statistically significant, ROS levels were increased in Atg5-deficient LSKs compared to controls, suggesting that impairment of autophagic clearance of damaged mitochondria causes functional defects of HSCs and results in disturbed differentiation of mature progenitors and terminally differentiated cells. Because ROS have normally harmful effect on cell survival, reduced apoptosis in Atg5-deficient LSKs seems paradoxical. However, as overall proliferation was also improved following Atg5 deficiency, we propose that this aberrant proliferation compensate the harmful effect of ROS that mediates apoptosis. Collectively, these findings demonstrate that Atg5 plays a critical role as a regulator to maintain HSCs and its reconstitution function, while future studies are needed to address the relationship between autophagy and other maintenance factors in HSCs.
 XML Download
XML Download