

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Various types of immune cells utilize specific cellular metabolic processes to support cell survival, development, and function (1). Metabolic reprogramming is crucial for T cell activation and function. In the steady state, naïve T cells use fatty acid oxidation for their basal metabolic needs. Upon the transition from naïve T cells to activated effector T cells, effector T cells rely on glycolysis, fatty acid synthesis, and amino acid metabolism to promote cell proliferation and cytokine secretion. In contrast to effector T cells, memory CD8+ T cells use the tricarboxylic acid cycle and fatty acid oxidation to promote cell longevity (234). Because the different stages of T cells use specific metabolic programs, understanding and modulating the proper metabolic process are necessary to elicit effective immune responses.
Bioenergetic drugs can be used to modulate the fate of T cells. 2-Deoxy-D-glucose (2-DG) is a glycolysis inhibitor that promotes CD8+ memory T cell formation (5). Rapamycin-mediated mTOR inhibition accelerates the memory CD8+ T cell response after viral infection (6). Although metformin-mediated AMP-activated protein kinase (AMPK) has been reported to enhance the memory CD8+ T cell response by modulating fatty acid metabolism after bacterial infection (7), deletion experiments showed that AMPK is dispensable for the primary immune response but necessary for the secondary immune response (8). However, no studies have evaluated the in vivo effects of AMPK activation on T cell differentiation after viral infection. A recent study indicated that constitutive glycolytic metabolism does not inhibit memory formation but promotes the differentiation of memory CD8+ T cells and effector-memory CD8+ T cells (9), suggesting that constitutively increased glycolysis generates sufficient ATP by T cells and induces a memory pool towards effector memory CD8+ T cells. However, the impact of a constitutive energy shortage in a metabolically restrictive environment on T cell differentiation has not been clearly demonstrated.
IM156 is a new bioenergetic biguanide derivative drug formerly known as HL156A. Similar to other biguanides, IM156 blocks mitochondrial complex I (1011). Studies have shown that after treatment of in vitro cultured rat peritoneal mesothelial cells and rat renal proximal tubular cells with IM156, AMPK activity is more potent than that with other AMPK agonists such as metformin or 5-aminoimidazole-4-carboxamide 1-β-D-ribofuranoside (1213). However, although IM156 treatment reduced the ATP levels in glioblastoma cell lines, AMPK activation by IM156 was not observed in these cell lines. This suggests that IM156 affects tumor cells via energy depletion caused by oxidative phosphorylation inhibition, but not because of an AMPK-dependent pathway (10). Taken together, these results suggest that IM156 treatment affects different modes of action depending on the cell type and often causes cellular metabolic perturbations and energy stress. However, the in vivo effects of IM156 on the differentiation and function of CD8+ T cells is unknown.
In this study, we investigated how IM156 treatment affects antigen-specific CD8+ T cell differentiation in vivo during acute infection with acute lymphocytic choriomeningitis virus (LCMV). We found that IM156 treatment increased the differentiation of memory CD8+ T cells in a dose-dependent manner, leading to impaired CD8+ T cell immune responses. Our results demonstrate that excessive AMPK activation by IM156 suppresses the differentiation and function of memory CD8+ T cells, suggesting that precise metabolic regulation is required to modulate T cell differentiation.
MATERIALS AND METHODS
Mice and viral infection
Five- to 6-wk-old female C57BL/6 mice were purchased from ORIENT BIO, Inc. (Seongnam, Korea). Mice were infected with 2×105 plaque-forming units of LCMV Armstrong (Arm) via intraperitoneal injection. All mice were maintained in a specific pathogen-free facility in accordance with Institutional Animal Care and Use Committee (IACUC) guidelines at Yonsei University. Animal experiments were approved by the IACUC of Yonsei University (201709-629-03).
Administration of IM156 and rapamycin to mice
From days −1 to 29 post-infection, IM156 (ImmunoMet Therapeutics, Inc., Houston, TX, USA) was intraperitoneally administered every other day at the indicated dose. Rapamycin (75 µg/kg; LC Laboratories, Wobum, MA, USA) was intraperitoneally administered daily. Control mice were administered daily injections of 5% DMSO during the treatment period.
Cell isolation, antibodies, and flow cytometry
PBMCs were isolated from the peripheral blood by Histopaque-1077 (Sigma-Aldrich, St. Louis, MO, USA) density gradient sedimentation. For phenotypic analysis of virus-specific CD8+ T cells derived from the peripheral blood and spleen, the cells were stained with the following fluorochrome-conjugated antibodies in phosphate-buffered saline containing 0.2% fetal bovine serum: antibodies against CD62L (MEL-14) and KLRG1 (2F1) (BD Biosciences, San Jose, CA, USA); antibodies against CD4 (RM4-5) (Biolegend, San Diego, CA, USA); and antibodies against CD8 (53-6.7) and CD127 (A7R34) (eBiosciences, San Diego, CA, USA) in the presence of a virus-specific tetramer. H-2Db tetramers bound to GP33-41 peptides were generated and used as previously described (14). For intracellular cytokine staining, splenocytes re-stimulated ex vivo with 0.2 µg/mL of LCMV GP33-41 peptide for CD8+ activation or GP66-80 peptide for CD4+ activation in the presence of brefeldin A (GolgiPlug; BD Biosciences) and monensin (GolgiStop; BD Biosciences) for 5 h. Stimulated cells were fixed, permeabilized, and stained with fluorochrome-conjugated antibodies against IL-2 (JE6-5H4), IFN-γ (XMG1.2), and TNF-α (MP6-XT22) (BD Biosciences). To remove the dead cell population, the Live/Dead Fixable Dead Cell Stain Kit (Invitrogen; Carlsbad, CA, USA) was used in the staining procedures. Flow cytometry was performed using the FACSCantoII (BD Biosciences). Data were analyzed using FlowJo software (TreeStar, Ashland, OR, USA).
RESULTS
IM156 inhibits the memory differentiation of antigen-specific CD8+ T cells
The in vivo effect of AMPK activation on T cell differentiation was evaluated using metformin in mice immunized with recombinant Listeria expressing ovalbumin (7). We tested the in vivo activity of IM156, a more potent AMPK activator than metformin, on CD8+ T cell differentiation during acute infection with LCMV Arm. We compared the effect of IM156 to that of rapamycin, a strong inhibitor of mTOR, because rapamycin treatment of LCMV-infected mice has been reported to enhance memory CD8+ T cell differentiation by inhibiting the glycolysis pathway in proliferating effector T cells. In this experiment, mice were infected with LCMV Arm and treated with IM156 or rapamycin. PBMCs were isolated from the mice on days 8, 15, and 30 post-infection to monitor the differentiation of antigen-specific CD8+ T cells. In contrast to tetramer+ T cells observed in CD8+ T cells, tetramer+ T cells were not detected in CD8-negative T cell populations. Similar to previous results (6), in the rapamycin-treated group, the frequency and number of virus-specific CD8+ T cells (GP33-tetramer positive CD8+ T cells) were increased during contraction and memory differentiation. However, the frequency and number of virus-specific T cells in IM156-treated mice did not change compared to in control mice. (Fig. 1A and B). To determine whether IM156 or rapamycin affects the differentiation of virus-specific CD8+ T cells, we measured kinetic changes by analyzing cell phenotypes. Most effector CD8+ T cells, referred to as short-lived effector cells (SLECs), are known to die because of apoptosis. In contrast, a small subset of cells referred to as memory precursor effector cells (MPECs) persists and differentiates into long-lived memory cells. Consistent with a previous report, rapamycin treatment increased the population of MPECs (CD127hiKLRG1lo) but decreased the population of SLECs (CD127loKLRG1hi) compared to control DMSO treatment. In contrast, IM156-treated mice showed significantly decreased populations of MPECs but increased populations of SLECs compared to control DMSO-treated mice (Fig. 1C and D). Memory T cells are divided into two subsets: central memory T cells (CD127+CD62L+) with a high proliferative potential, which located in secondary lymphoid organs, and effector memory T cells (CD127+CD62L−) with a rapid effector function and cytotoxicity, which circulate through peripheral organs (15). Particularly, the frequency of effector memory T cells was dramatically reduced in IM156-treated mice (Fig. 1C and E). These results suggest that IM156 inhibits the memory differentiation of antigen-specific CD8+ T cells.
 | Figure 1IM156 reduces antigen-specific effector memory T cells in LCMV-infected mice.LCMV-infected mice were treated with DMSO (
 ), IM156 ( ), IM156 ( ), or rapamycin ( ), or rapamycin ( ) from days −1 to 29 post-infection. (A) Representative plot showing the frequency of GP33 tetramer-positive cells among CD8+ or CD4+ T cells in PBMCs at the indicated days post-infection. (B) Kinetics of GP33+ T cells among CD8+ T cells as in A. (C) Co-expression of either KLRG1 (left) or CD62L (right) and CD127 on tetramer-positive CD8+ T cells obtained from PBMCs at the indicated days post-infection. (D) Frequency of MPECs (CD127hiCD62Llo) and SLECs (CD127loCD62Lhi) as in C. (E) Frequency of TCM (CD127+CD62L+), TEM (CD127+CD62L−), and TEFF (CD127−CD62L−) as in C. Data are representative of 3 independent experiments with n=4 mice per group. Results are the mean ± SEM and statistical significance was determined by 2-tailed unpaired Student's t-test. ) from days −1 to 29 post-infection. (A) Representative plot showing the frequency of GP33 tetramer-positive cells among CD8+ or CD4+ T cells in PBMCs at the indicated days post-infection. (B) Kinetics of GP33+ T cells among CD8+ T cells as in A. (C) Co-expression of either KLRG1 (left) or CD62L (right) and CD127 on tetramer-positive CD8+ T cells obtained from PBMCs at the indicated days post-infection. (D) Frequency of MPECs (CD127hiCD62Llo) and SLECs (CD127loCD62Lhi) as in C. (E) Frequency of TCM (CD127+CD62L+), TEM (CD127+CD62L−), and TEFF (CD127−CD62L−) as in C. Data are representative of 3 independent experiments with n=4 mice per group. Results are the mean ± SEM and statistical significance was determined by 2-tailed unpaired Student's t-test.
*p<0.05; **p<0.01; ***p<0.001.
|
IM156 delays conversion of CD127 and CD62L of antigen-specific CD8+ T cell in a dose-dependent manner
To investigate the optimal in vivo dose of IM156 for modulating the virus-specific CD8+ T cell response, LCMV virus-infected mice were treated with 1, 5, or 20 mg/kg IM156. The frequencies of GP33 tetramer-positive cells among CD8+ T cells in the blood and spleen did not differ among the groups of mice treated with different doses of IM156 (Figs. 2A, 2B, 3A, and 3B). To analyze the phenotype of memory CD8+ T cells in the blood of IM156-treated mice during their differentiation, we performed phenotypic analysis of virus-specific CD8+ T cells to determine the expression of CD127, CD62L, and KLRG1. Although both CD62L and KLRG1 expression on virus-specific CD8+ T cells did not differ in the group of mice treated with different doses of IM156 at days 8 and 15 post-infection, CD127 expression was significantly decreased depending on the dose of IM156 even at day 15 post-infection (Fig. 2C and D). At day 30 post-infection, the IM156 dosage effect was shown to be prominently reflected as the delayed conversion of both CD127 and CD62L (Fig. 2C and D).
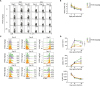 | Figure 2IM156 dose-dependently impairs CD127 conversion of antigen-specific CD8+ T cells.LCMV-infected mice were treated with various concentrations of IM156 from days −1 to 29 post-infection. (A) Representative plot showing the frequency of GP33 tetramer-positive cells among CD8+ or CD4+ T cells in PBMCs of IM156-treated mice at the indicated days post-infection. (B) Frequency of GP33+CD8+ T cells as in A. (C) Expression levels of CD127 (left), CD62L (middle), and KLRG1 (right) on tetramer-positive CD8+ T cells in PBMCs of IM156-treated mice at the indicated time post-infection. The relative percentages of CD127, CD62L, and KRLG1-expressing cells are indicated. (D) MFI of CD127 (top), CD62L (middle), and KLRG1 (bottom) on virus-specific CD8+ T cells in PBMCs as in C.
Data are representative of 3 independent experiments n=4–5 mice per group. Results are the mean ± SEM and statistical significance was determined by two-tailed unpaired Student's t-test.
*p<0.05; **p<0.01; ***p<0.001.
|
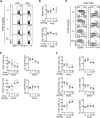 | Figure 3IM156 dose-dependently suppresses the differentiation of central memory T cells and effector memory T cells.LCMV-infected mice were treated with various concentrations of IM156 from days −1 to 29 post-infection. (A) Representative data showing the frequency of GP33 tetramer-positive cells among CD8+ or CD4+ T cells in the spleen at day 30 post-infection. (B) Frequency (top) and absolute number (bottom) of GP33+CD8+ T cells as in A. (C) Co-expression of either KLRG1 (left) or CD62L (right) and CD127 on tetramer-positive CD8+ T cells obtained from the spleen at day 30 post-infection. (D) Frequency (left) and absolute number (right) of MPECs (CD127hi KLRG1lo) and SLECs (CD127loKLRG1hi) as in C. (E) Frequency (left) and absolute number (right) of TCM (CD127+CD62L+), TEM (CD127+CD62L−), and TEFF (CD127−CD62L−) as in C. Data are representative of 3 independent experiments n=4–5 mice per group. Results are the mean ± SEM and statistical significance was determined by 2-tailed unpaired Student's t-test.
*p<0.05; **p<0.01; ***p<0.001.
|
IM156 dose-dependently increases SLECs but decreases MPECs
The frequencies and absolute numbers of MPECs in the spleen of IM156-treated mice also decreased in a dose-dependent manner at day 30 post-infection (Fig. 3C and D). Notably, both central memory T cells and effector memory T cells were decreased in the spleen of IM156-treated mice in a dose-dependent manner (Fig. 3C and E). Particularly, treatment with 20 mg/kg of IM156 strongly inhibited the differentiation of virus-specific CD8+ T cells into memory cells.
We analyzed the expression of KRLG1 and CD62L versus CD127 gated on total splenic CD4+ T cells. Although they are not antigen-specific CD4+ T cells, CD127+CD62L+ cells were slightly decreased among total CD4+ T cells by 5 mg/kg IM156 (Supplementary Fig. 1). This suggests that IM156 slightly affects the composition of CD4+ T cells, the differentiation of antigen-specific CD4+ T cells remains unknown.
IM156 impairs the ability of antigen-specific CD8+ T cells to produce cytokines
To examine whether in vivo treatment of IM156 affects the function of memory T cells, splenocytes obtained at day 30 post-infection were re-stimulated ex vivo with the virus-specific peptide GP33-41 for CD8+ T cells and GP66-80 for CD4+ T cells. The percentage and expression levels of TNF-α- and IL-2-producing cells among IFN-γ+CD8+ T cells decreased dose-dependently in the IM156-treated group (Fig. 4A and B). Similar results were observed for IFN-γ+CD4+ T cells in IM156-treated mice (Fig. 4C and D). It has been reported that CD107a represents cytotoxic CD8+ and CD4+ T cell responses to viral and tumor antigen associated with T cell cytolytic potential (16171819). However, there was no difference in CD107a expression on CD8+ and CD4+ T cells after in vivo treatment with various doses of IM156, indicating that IM156 does not affect the cytotoxicity of T cells (Supplementary Fig. 2). Although the cytotoxic potential was not altered by IM156, the polyfunctionality of CD8+ and CD4+ T cells was significantly decreased after IM156 treatment as shown in Fig. 4, suggesting that IM156 modulates the ability of T cells to produce effector cytokines but not their cytotoxicity after viral infection.
 | Figure 4IM156 inhibits multi-functional CD8+ and CD4+ T cells.LCMV-infected mice were treated with various concentrations of IM156 from days −1 to 29 post-infection. (A) Representative flow cytometric analysis of production of IFN-γ, TNF-α, and IL-2 in CD8+ T cells obtained from the spleen at day 30 post-infection after ex vivo re-stimulation with GP33 peptide. (B) Frequency of TNF-α-producing cells (top left) and IL-2-producing cells (top right) among IFN-γ+CD8+ T cells. MFI of TNF-α (bottom left) and IL-2 (bottom right) among IFN-γ+CD8+ T cells as in A. (C) Representative flow cytometric analysis of production of IFN-γ, TNF-α, and IL-2 in CD4+ T cells obtained from the spleen at day 30 post-infection after ex vivo re-stimulation with GP66 peptide. (D) Frequency of TNF-α-producing cells (top left) and IL-2-producing cells (top right) among IFN-γ+CD4+ T cells. MFI of TNF-α (bottom left) and IL-2 (bottom right) among IFN-γ+CD4+ T cells as in C. Data are representative of 3 independent experiments n=4–5 mice per group. Results are the mean ± SEM and statistical significance was determined by 2-tailed unpaired Student's t-test.
*p<0.05; **p<0.01; ***p<0.001.
|
Taken together, our data indicate that IM156 treatment delays the differentiation of antigen-specific CD8+ and CD4+ T cells into memory cells, resulting in an incomplete functional capacity to produce effector cytokines upon the exposure to the same antigen.
DISCUSSION
The objective of this study was to investigate the in vivo effects of IM156, a potent AMPK activator, on the differentiation of antigen-specific CD8+ T cells after acute virus infection. Our study showed that IM156 inhibited the differentiation and function of memory CD8+ T cells in a dose-dependent manner. The conversion rate of CD127 molecules on antigen-specific CD8+ T cells in the blood was significantly decreased by IM156 treatment (Fig. 2). This is the first study to determine the in vivo role of IM156 in regulating antigen-specific CD8+ T cell differentiation.
While metformin-mediated AMPK activation enhances memory T cell differentiation (7), APMKα1 deletion leads to reduced CD8+ memory T cell function during bacterial infection (8). One of the consequences of AMPK activation is inhibition of the mTOR pathway. Consistent with our results, the mTOR inhibitor rapamycin promoted the generation of memory T cells (6), suggesting that inhibition of glycolytic metabolism enhances the differentiation of memory T cells. The discrepancy in the effects caused by either metformin or IM156 may be related to the strength of AMPK activation and ability of cell penetration (1220). In a previous study, the mode of action of IM156 was described as activation of AMPK (111213) and/or energy deprivation as a result of inhibition of mitochondrial oxidative phosphorylation (1011). The best-known mechanism of IM156 is AMPK activation. IM156-mediated AMPK activation ameliorates peritoneal fibrosis (12), liver fibrosis (20), and renal fibrosis (13) and prevents the growth of human oral cancer (11). In our study, we showed that IM156 restricts proper memory differentiation after viral infection. Given that IM156 is a much more potent activator of AMPK than other biguanides including metformin, our data suggest that adequate AMPK activity is beneficial for generating memory T cells, while excessive AMPK activity can inhibit the differentiation of memory T cells. Another possibility for explaining this discrepancy between metformin and IM156 is the difference in the levels of energy deprivation when the drugs were targeted to antigen-specific T cells. IM156-mediated inhibition of mitochondrial function often prevents the growth of human oral cancer (11) and glioblastoma (10). Although the precise action mechanism of IM156 on virus-specific CD8+ T cells after viral infection remains unclear, IM156 likely also prevents the growth of memory progeny by inhibiting mitochondrial function.
A previous study showed that IM156 inhibited lipopolysaccharide-induced macrophage inflammation (21); however, it remains unclear whether the changes in T cell differentiation by IM156 are directly mediated by T cells or indirectly mediated by other cells. Because the T cell differentiation status according to AMPK activation is more important in the secondary immune response than in the primary immune response (8), the effect of IM156 during the recall response should be evaluated to test this impaired ability in secondary infection.
In summary, we identified the in vivo role of IM156, a potent AMPK activator, in determining the fate and function of memory T cells. Additionally, our results provide a potential strategy for suppressing memory T cell responses by using IM156. In lupus models, the combination of metformin and 2-DG normalized CD4+ T cell metabolism and showed therapeutic effects in lupus-prone mice (22). We suggest that IM156 is a potent AMPK activator that may have broader applications in suppressing auto-reactive memory T cells against self-antigens in autoimmune diseases.
 XML Download
XML Download