

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
RNA editing is a unique post-transcriptional modification that makes a mature RNA sequence different from its template DNA sequence by inserting, deleting, or substituting bases [123]. Because RNA editing alters the cellular fate of RNA molecules and their sequence relative to the genome, it plays a critical role in many biological processes [456789]. In mammals, the most common forms of RNA editing are adenosine deamination to inosine and cytidine deamination to uridine (C-to-U). Deficiency in any of the forms of RNA editing can lead to physiological defects or metabolic disorders [456789].
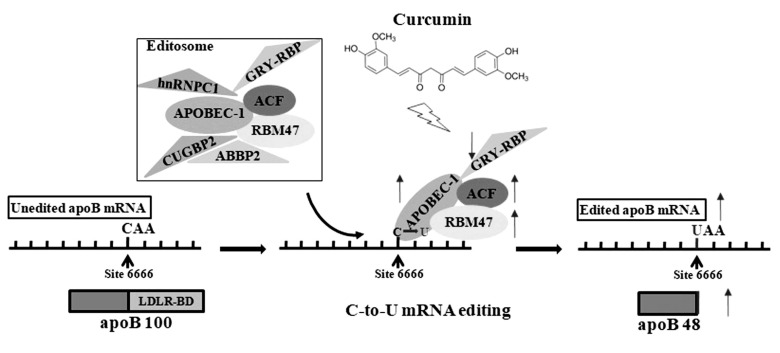
The best characterized C-to-U editing event in mammals occurs in the apolipoprotein B (apoB) mRNA. The C residue at position 6666 of the apoB mRNA is substituted with a U that changes a glutamine codon (CAA) to a termination codon (UAA), generating a truncated protein, apoB-48 [10]. This editing event has important effects on lipoprotein metabolism because apoB plays a central role in the assembly, secretion, and metabolism of low-density lipoproteins (LDLs). ApoB mRNA translates to apoB-100 protein in the absence of C6666RNA editing, and is virtually the only protein component of LDLs that contains the low-density lipoprotein receptor-binding domain; it binds with low-density lipoprotein receptor (LDLR), and mediates the metabolism of LDLs [10]. Due to C6666RNA editing, apoB-48 has only the N-terminal half of apoB-100, and lacks the domain that mediates interaction with the LDLR. Therefore, lipoprotein particles containing apoB-48 are directed to a different metabolic pathway. They possess multiple copies of apolipoprotein E, which assists in the clearance of lipoprotein, through LDLR and LDL receptor-related proteins [1112]. Actually, lipoprotein particles containing apoB-48 clear from plasma more rapidly than those containing apoB-100 (10 min compared to 3 days) [13]. In theory, if it were possible to manipulate or mimic apoB mRNA 6666 C-to-U editing, the secretion of lipoproteins containing apoB-48 could be upregulated, lipoproteins would undergo rapid catabolic clearance, thus reducing the risk of lipid metabolism disorders [14].
Curcumin, an extract from the traditional Chinese medicinal herb Curcuma longa L., possesses numerous important biological activities, including hypocholesterolemic properties [15]. A number of studies have shown that curcumin significantly reduces plasma LDL-cholesterol and improves lipid metabolic disorders [1617]. Liang et al.[18] found that curcumin could significantly reduce the expression of apoB-100 mRNA in live nonalcoholic fatty liver disease mouse, thereby regulating their lipid metabolism. In our previous study, we found that 15 M curcumin raised the level of apoB-48 and reduced the level of apoB-100 by increasing the expression of apoB editing catalytic polypeptide-1 (APOBEC-1) in primary rat hepatocytes [19]. However, apoB mRNA editing is mediated by a coordinated modulation of multiple enzyme components that includes not only APOBEC-1, but also apobec-1 complementation factor (ACF) [20] and RNA-binding-motif-protein-47 (RBM47) [21]. Other auxiliary proteins, including CUG binding protein-2 (CUGBP2), glycine-arginine-tyrosine-rich RNA binding protein (GRY-RBP), heterogenous nuclear ribonucleoprotein (hnRNP)-C1, and apobec-1 binding protein-2 (ABBP2), also participate in this editing event [22]. In this study, we used primary mouse hepatocytes as a model system to investigate the change in APOBEC-1, ACF, RBM47, CUGBP2, GRY-RBP, hnRNP-C1, and ABBP2 expression levels after treatment with curcumin to further clarify the specific functional role of each editing enzyme component in the apoB mRNA editing event.
METHODS
Isolation of primary hepatocytes
Institute of Cancer Research (ICR) mice, within 24 h of birth, were purchased from Shanghai Snooker Company (Shanghai, China). Animal experimental procedures were performed in conformity with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health publication no. 85-23, revised 1996) and were approved by the Ethics Committee of the Animal Laboratory Research Centre of Zhejiang Chinese Medical University (Zhejiang, China; approval no. SYXK [Zhe] 2008-0115). Primary hepatocytes were isolated from ICR mice livers as follows: ICR mice were decapitated and the liver tissues were extracted and placed in cold D-Hanks solution. Liver capsules were peeled off with forceps, and the livers were shredded directly with ophthalmic scissors. Tissues were thoroughly washed and then digested with 0.2% trypsin containing ethylenediaminetetraacetic acid at 37℃ with intermittent shaking. After discarding the supernatant, Dulbecco's modified Eagle's media (DMEM) containing 0.2% type II collagenase was added followed by digestion for 20 min at 37℃. The reaction was terminated by adding DMEM supplemented with 10% fetal bovine serum (FBS); the dispersed cells were filtered through a 200-mesh sieve and centrifuged for 5 min at 800 g. The isolated hepatocytes were washed twice with fresh DMEM and incubated at 37℃ in a humidified atmosphere of 5% CO2. After removing the cultured impurities, the first to third generation cells were used for the subsequent experiments. Cell viability, determined using trypan blue staining, was > 95%.
Identification of primary hepatocytes by periodic-acid-Schiff (PAS) reaction
Primary hepatocytes were seeded in 25 cm2 culture flasks and cultured in DMEM supplemented with 10% FBS. Following a 48 h incubation, the medium was replenished every 24 h, hereafter. Cells (5 × 104 cells/ml) were plated onto glass cover slips in 24-well culture plates and grown for 24 h. They were washed three times with phosphate buffer saline (PBS; pH 7.4), then fixed with fixation solution for 10 min, and permeabilized with 0.1% TritonX-100 for 20 min. Thereafter, the cells were stained using a periodic acid-Schiff kit according to the manufacturer's instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Finally, the stained cells were mounted on glass slides and photographed using a Nikon Eclipse 80i microscope connected to a DS-5M-L1 (Nikon, Melville, NY, USA) camera.
MTT-based cell viability assay
The effect of curcumin on cell viability was determined by MTT assay. Briefly, when the cells reached sub-confluence, they were seeded at a density of 5 × 103 cells/well in 96-well plate and treated with increasing doses of curcumin prepared in DMEM containing 10% FBS. MTT solution (0.5 mg/ml) was added to each well and incubated for 4 h at 37℃. The medium was removed; formazan crystals formed inside the cells were dissolved in 150 µl dimethyl sulfoxide, and then gently shaken for 15 min. The absorbance of MTT-formazan was measured at 570 nm using a SpectraMaxM3 microplate reader (MolecularDevices, Sunnyvale, CA, USA).
Real-time quantitative polymerase chain reaction (qRT-PCR)
Total RNA was isolated from the primary hepatocytes using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according to the manufacturer's instructions. Equal quantities (5.0 µg) of total RNA were used for complementary DNA (cDNA) synthesis using the HiFi-MMLV cDNA kits (Beijing Kang Century Biotechnology Co., Ltd., Beijing, China).Ultra SYBR mixture (Beijing Kang Century Biotechnology Co.,Ltd.) was used to perform qRT-PCR. Relative expression of the target gene was normalized against β-actin and analyzed via the 2−ΔΔCt method. All PCR-specific forward and reverse primers were synthesized by GenScript Co., Ltd. (Nanjing, China) (Table 1).
Western blot analysis
Primary hepatocytes were washed once with PBS and dissolved in Protein Extraction Reagent (Boster Bioengineering, Wuhan, China) containing 1 mM phenylmethanesulfonyl fluoride (PMSF; Roche Molecular Biochemicals, Indianapolis, IN, USA). Total protein was electrophoretically separated with sodium dodecyl sulfate-polyacrylamide gel electrophoresis, followed by transfer to polyvinylidene difluoride (PVDF) membrane. The PVDF membrane was blocked with 1% bovine serum albumin for 2 h at room temperature, and then incubated with the antibodies, separately, overnight at 4℃. The antibody against β-actin was purchased from Hua Bio (Hangzhou HuaAn Biotechnology Co., Ltd., Hangzhou, China), antibodies against APOBEC-1, CUGBP2, and (hnRNP)-C1 were from Santa Cruz (Santa Cruz Biotechnology, Dallas, TX, USA), antibody against ACF (Thermo Fisher Scientific, Waltham, MA, USA), RBM47, and ABBP2 (Proteintech Group, Inc., Chicago, IL, USA), GRY-RBP (Abcam, Cambridge, UK). The membrane was then washed with tris buffered saline-tween solution three times followed by incubation with anti-rabbit immunoglobulin G (IgG) or anti-mouse IgG horseradish peroxidase secondary antibody (Hangzhou HuaAn Biotechnology Co., Ltd.) for 2 h at room temperature. Finally, the protein bands were detected by enhanced chemiluminescence (ECL) detection kit (Amersham Biosciences, Piscataway, NJ, USA). Relative protein expression levels were quantified by using Image J software and normalized to β-actin.
Quantification of C-to-U RNA editing level by sequencing
Sequencing of apoBC6666 sites was performed by the direct sequencing method to estimate the editing frequency (Shanghai Sangon Biological Engineering Technology & Services Co., Ltd., Shanghai, China). Briefly, cDNA from three independent biological replicates was PCR amplified using Taq DNA Polymerase (TaKaRa, Otsu, Japan); the primers are listed in Table 1. The PCR product was purified using the AxyPrep PCR cleanup Kit (Axygen Scientific Inc., Union City, CA, USA) and sequenced using the forward primer. In parallel, we also extracted the hepatocyte genome and PCR amplified apoB gene fragments as a control. The ratio between the C and T peak height in individual chromatograms was measured using FinchTV (https://digitalworldbiology.com/FinchTV). Percent editing was calculated as the T peak height relative to the total peak height [T / (C + T) × 100].
siRNA-mediated gene-specific knockdown
Lipofectamine 2000 (Invitrogen, Life Technologies, Carlsbad, CA, USA) was used to introduce APOBEC-1 siRNA (MSS202127), ACF siRNA (MSS245486), and RBM47 siRNA (MSS205172) oligonucleotides (Invitrogen, Life Technologies) into primary hepatocytes according to the manufacturer's instructions. A scrambled siRNA was used as the negative control. After 48 h incubation, total RNA was isolated from the transfected primary hepatocytes and qRT-PCR was performed using specific primers (Table 1) to evaluate the effect of siRNA-mediated gene knockdown.
Statistical analysis
Data are expressed as mean ± standard deviation. Statistical analysis was performed using one-way analysis of variance for multiple comparisons and t-tests among groups. The data were analyzed using IBM SPSS Statistics 21.0 (IBM, Armonk, NY, USA). Statistical significance was considered if p < 0.05.
RESULTS
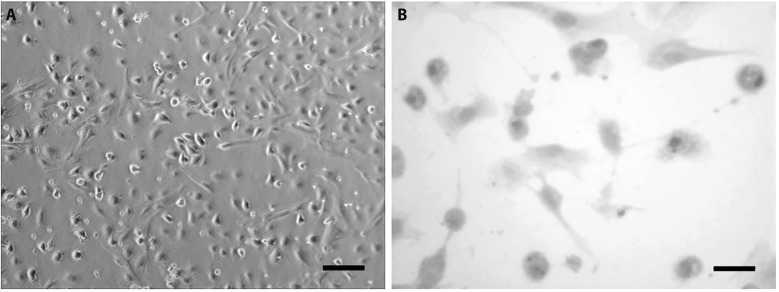
Identification of primary mouse hepatocytes by PAS staining
Hepatocytes, isolated immediately from newborn mouse liver had a rounded cell body with a translucent cytoplasm. After overnight incubation, the cells appeared more polygonal in shape (Fig. 1A). Healthy hepatocytes were devoid of granulations or blebs. To identify the hepatocytes, we performed periodic acid-Schiff staining. As shown in Fig. 1B, dense pink glycogen granules were observed in the cytoplasm of the cells, around the nuclei.
Effects of curcumin on the viability of primary mouse hepatocytes
The MTT assay was used to determine the dose range of curcumin that does not affect the proliferation of primary mouse hepatocytes. As shown in Fig. 2A, the cell viability was not significantly repressed by treatment with < 50 µM dose of curcumin for 24 h, but treatment with 60 µM curcumin resulted in an approximate 40% decrease in cell survival. Therefore, three concentrations (5, 25, and 50 µM) were selected to further assess the cytotoxicity of curcumin after 36 h and 48 h of treatment. Results showed no decrease in viability between the control (0 µM) and curcumin-treated (5, 25, and 50 µM) cells after 36 h and 48 h (Fig. 2B). Therefore, 5, 25, and 50 µM curcumin were considered the optimum concentration for subsequent experiments.
Curcumin increases the efficiency of apoB mRNA editing
To determine how curcumin affects the ability of primary mouse hepatocytes to edit apoB mRNA, total cellular RNA was isolated from primary hepatocytes after treatment with 5, 25, and 50 µM curcumin for 48 h and then assayed for apoB mRNA editing efficiency as described in materials and methods. As shown in Fig. 3, the C-to-U editing signal was detected in all the RT-PCR sequencing groups, but not in the genome which was isolated from 50 µM curcumin treated hepatocytes. The editing efficiency in isolated hepatocytes varied with each individual mouse and ranged from 5.13%–8.05% (n = 7); after treatment with 5 and 25 µM curcumin, the editing efficiency demonstrated no significant changes (4.48%–8.62%, 5.06%–8.41%, respectively). However, increasing the concentration of curcumin to 50 µM, enhanced the editing efficiency of apoB mRNA C6666 site (27.63%–35.61%), indicating that treatment with an intermediate concentration of curcumin increased the amount of apoB-48 and reduced the amount of apoB-100 in cultured primary mouse hepatocytes.
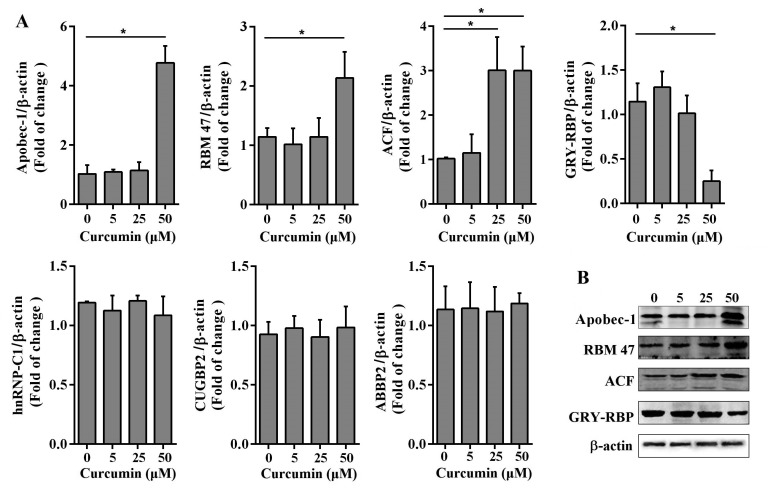
Effect of curcumin on apoB mRNA editing enzyme components
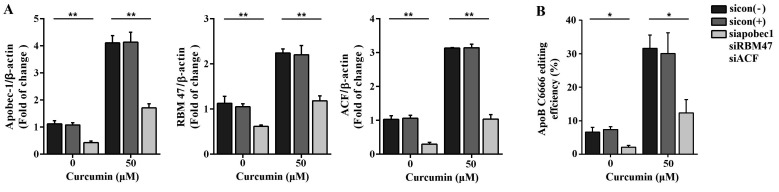
To further explore the mechanism of curcumin in upregulating apoB mRNA editing, we evaluated the change in the expression of the multiple enzyme components following curcumin treatment, by qRT-PCR and western blotting. As shown in Fig. 4A, the mRNA level of APOBEC-1 was approximately four-fold higher in cells treated with 50 µM curcumin than in untreated, 5, and 25 µM curcumin-treated cells. The mRNA levels of RBM47 and ACF were also upregulated by 50 µM curcumin treatment. Interestingly, 25 µM curcumin treatment increased the mRNA level of ACF as well. Furthermore, we found that treatment with 50 µM curcumin significantly reduced the mRNA level of GRY-RBP, an inhibitory component of C-to-U RNA editing enzyme complex, while curcumin treatment had no effect on other auxiliary proteins CUGBP2 and hnRNP-C1. In addition, protein expression was consistent with the mRNA status (Fig. 4B). Next, APOBEC-1, ACF, and RBM47 siRNA oligonucleotides were transfected into primary hepatocytes to evaluate whether the increased apoB RNA editing by 50 µM curcumin disappeared when the expression of the editosome components was knockdowned. As shown in Fig. 5A, both in untreated and 50 µM curcumin treated group, the expression of APOBEC-1, ACF, and RBM47 was decreased in siRNA-mediated gene knockdown cells compared with siRNA NC transfected and untransfected cells. As anticipated, the editing efficiency of apoB mRNA C6666 site in APOBEC-1, ACF, and RBM47 knockdown hepatocytes was significantly decreased, only had 1.47%–2.62% (Fig. 5B). In 50 µM curcumin treated group, the editing efficiency in APOBEC-1, ACF, and RBM47 knockdown hepatocytes was also reduced (8.43%–16.27%) compared with siRNA NC transfected and untransfected hepatocytes (Fig. 5B). Collectively, these results suggested that curcumin affected apoB mRNA editing efficiency by regulating the expression of the components of the editing enzyme complex.
DISCUSSION
ApoB mRNA editing is an exquisitely precise nuclear process that alters a single nucleotide in a molecule containing over 14,000 bases. This editing event requires the assembly of a multiprotein editosome. Among them, APOBEC-1, which has cytidine deaminase activity, is the catalytic component of the editing enzyme complex [23]. ACF (recently renamed A1CF) is an RNA-binding protein that serves as the RNA recognition component of the editing enzyme [2425], while RBM47 is also involved in the editing events via interaction with APOBEC-1 and ACF [21]. Besides the afore mentioned core components, the editosome also includes other auxiliary inhibitory proteins such as CUGBP2, GRY-RBP, hnRNP-C1, and ABBP2 [22262728].
Studies have indicated that apoB mRNA editing is tissue-specific [293031] and modulated by several interventions (e.g. insulin, ethanol), which could regulate the expression of APOBEC-1 or the auxiliary proteins [323334353637]. Because there are differences in the apoB100- and apoB48-containing nascent particles in their dependence on a second step of lipidation [38], and the rate of clearance of their respective remnants from the circulation, the degree of apoB mRNA editing has important physiological implications.
Curcumin has been shown to regulate the lipid mechanism in hepatocytes [3940] as well as the lipoprotein concentrations in serum [4142]. Our previous study showed that intermediate concentration of curcumin increased the degree of apoB mRNA editing in primary murine hepatocytes by up-regulating the expression of APOBEC-1. The current study aimed to clarify the effect of curcumin on other components of editosome. As shown in Fig. 4, in primary mouse hepatocytes, 50 µM curcumin significantly increased the mRNA and protein levels of APOBEC-1, ACF, and RBM47, but decreased the levels of GRY-RBP. No effect of curcumin was observed on inhibitory proteins CUGBP2, hnRNP-C1, and ABBP2. In addition, the increased apoB RNA editing by 50 µM curcumin was significantly reduced by siRNA-mediated knockdown of APOBEC-1, ACF, and RBM47. These results suggested that the intermediate concentration of curcumin enhances the apoB mRNA editing efficiency by regulating the expression of all the three core components, but not the auxiliary inhibitory members in editing enzyme complex (Fig. 6); there is a coordinated modulation among these four components in vivo. The increased, and similar expression of ACF at 25 µM and 50 µM curcumin, suggests that curcumin does not affect apoB mRNA editing in a dose-dependent manner. Researches have shown that curcumin increased the expression of CUGBP2 in pancreatic cancer cells and inhibited pancreatic tumor growth via the CUGBP2-mediated post-transcriptional regulation pathway [4344]. However, we didn't detect the influence of 5, 25, and 50 µM curcumin on the mRNA level of CUGBP2 in primary mouse hepatocytes. Curcumin has been reported to possess antitumor properties but does not cause any damage to the normal cells, and downregulated the expression of bcl-2 in two human breast adenocarcinoma cell lines but had no significant effect on bcl-2 expression in normal rat hepatocytes [45]. So we believed that the effect of curcumin on gene expression in normal cells was different with that in malignant cells.
Van Mater et al. [46] reported that apoB mRNA editing efficiency in isolated, primary rat hepatocytes ranged from 38% to 58% and varied with each individual rat, but the proportion of edited apoB mRNA in our primary mouse hepatocytes was much lower, ranging from 5.13% to 8.05%. It is worth noting that Van Mater et al. [46] used hepatocytes at the time of their preparation, before they were plated. In our study, primary hepatocytes were treated with curcumin for 48 h, and when they were used to assay for apoB mRNA editing efficiency, it was more than 96 h from the time of their isolation. In addition, Van Mater et al. [46] isolated hepatocytes from rat, and we isolated hepatocytes from mouse. Accordingly, we considered that the differences in the data may be due to tissue-specific expressions and nutritional perturbations.
Unlike the dramatic change in apoB mRNA editing efficiency that is apparent with adenoviral delivery of exogenous APOBEC-1 [47], curcumin-induced changes in the apoB mRNA editing efficiency were comparatively small, because curcumininduced editing events involved the coordinated expression of multiple editing components in vivo. In conclusion, our findings throw a new light on the hypolipidemic effect of curcumin that elevates the level of apoB-48 and reduces the level of apoB-100 by coordinating the expression of some components of the editosome; curcumin may be used therapeutically to reduce levels of atherogenic lipoproteins and prevent atherosclerosis.



 XML Download
XML Download