

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Obesity is a major risk of serious diet-related chronic diseases, such as hyperlipidemia, hypertension, type II diabetes, atherosclerosis, cardiovascular disease, non-alcoholic fatty liver disease, and metabolic syndrome [123]. These obesity-related syndromes are caused by triglyceride (TG) overload and fat deposition [45]. When TG levels are excessive, TG overload will cause lipid accumulation and interfere with the normal function of adipocytes. Excess fat deposition is causally linked to metabolic syndrome abnormalities [67]. Some studies have suggested that adipogenesis and lipolysis were considered as the complex processes involving several transcription factors, such as lipogenesis genes (the peroxisome proliferator-activated receptors [PPAR-α, PPAR-β, and PPAR-γ], CCAAT/enhancer-binding proteins [C/EBP-α and C/EBP-β], and sterol regulatory element-binding proteins [SREBP-1a, SREPB-1c, and SREBP-2]) [89], lipid synthesis enzymes (the fatty acid-binding protein [aP2], lipoprotein lipase [LPL], and fatty acid synthase [FAS]) [101112], and lipolysis genes (the hormone-sensitive lipase [HSL], aquaporin-7 [AQP-7], and adipose triglyceride lipase [ATGL]) [131415]. Hence, blocking the expressions of lipid metabolism-related transcription factors and their productions would be an impactful way to ameliorate lipid accumulation and improve obesity-related chronic diseases.
Long-term dietary supplementation intervention is important for humans at risk of developing adiposity [1617]. Teas have long been considered as a dietary component [18]. Rhizophora stylosa (Rhizophoraceae) is identified as a welcome edible plant whose leaves and roots are commonly used as an ingredient for herbal tea in Southern China [19]. As shown in Fig. 1A, fumigaclavine C (FC), an active indole alkaloid, is obtained from endophytic Aspergillus terreus (strain No. FC118) by the root of Rhizophora stylosa (Rhizophoraceae). FC possesses multiple health beneficial effects, including anti-inflammation [20], anti-atherosclerosis [21], anti-tumor [22], hepatoprotective activity [23], and immunosuppressive activity [24]. A previous study indicated that FC had a potential anti-atherosclerosis activity in apolipoprotein E-deficient mice via activating PPARs signaling pathway [25]. In high-fat diet (HFD)-induced obese animal model, the fat intake of obesity is accompanied with a low-grade inflammation characteristic [262728]. However, the anti-obesity effect of FC and the precise molecular mechanisms are incompletely delineated. Hence, this study is designed to evaluate whether FC has anti-adipogenic effect in adipocytes and whether it improves lipid accumulation in HFD-induced obese mice.
METHODS
Materials
FC (purity is 99.5% by high performance liquid chromatography) is obtained from endophytic Aspergillus terreus (strain No. FC118) by the root of Rhizophora stylosa (Rhizophoraceae). The primary antibodies including PPAR-α, PPAR-β, PPAR-γ, C/EBP-α, C/EBP-β, SREBP-1c, aP2, LPL, FAS, HSL, AQP-7, ATGL, β-actin, and alkaline phosphatase labeled secondary antibody were purchased from Cell Signaling Technology (Danvers, MA, USA). Glycerol, TG, total cholesterol (TC), and Cell Counting Kit-8 (CCK-8) were evaluated via diagnostic assay kits from Nanjing Jiancheng Company (Nanjing, China). Simvastatin (Sim) was purchased from Sigma Aldrich Company (St. Louis, MO, USA).
Cell culture and differentiation
3T3-L1 pre-adipocyte cells were purchased from American Type Culture Collection (ATCC, Rockville, MD, USA). The cells were cultured in dulbelcos modified eagles medium (DMEM) containing heat-inactivated fetal calf serum (10%), streptomycin (100 µg/ml), and penicillin (100 U/ml) at 37℃ with 5% CO2. 3T3-L1 adipocyte cells viability was evaluated by CCK-8 diagnostic assay kit following the manufacture's specification. The cells were stimulated with 1-isobutyl-3-methylxanthine (0.5 mM), dexamethasone (1 µM), and insulin (10 µg/ml) in 6-well plates for 48 h. The medium was then replaced with DMEM containing insulin (10 µg/ml) for 48 h. These cells were incubated in DMEM without insulin every 48 h until 96 h. FC was dissolved in DMSO (final concentration ≤ 0.1%). The differentiated 3T3-L1 adipocyte cells were treated with various concentrations of FC for 24 h. Cell supernatants, proteins, and RNA extracts were stored in −80℃ refrigerator. Treatment with Sim (10 µM) was used as a positive control.
Animals and experimental design
Four-week-old male C57BL/6 mice were purchased from SJA Laboratory Animal Co., Ltd. (Hunan, China), and were acclimatized to the experimental facility for one week. The mice were kept in a temperature and humidity controlled room with ad libitum access to water and mouse chow diet (CD, 10% fat, 14% protein, and 76% carbohydrates, 3.6 kcal/g) or high fat diet (HFD, 50% fat, 14% protein, and 36% carbohydrates, 5.1 kcal/g [Shanghai FBSH Biological Pharmaceutical Co., Ltd., Shanghai, China]) for sixteen weeks. All experimental procedures and care administered were approved by the Institutional Animal Care and Use Committee of Liuzhou General Hospital (Protocol No. 1019-3). Experiments with animals were performed following the animal ethics guidelines of the Institutional Animal Ethics Committee. All animals were randomly subdivided into five groups with ten mice in each group who were treated as follows for fifteen weeks. FC (10 and 20 mg/kg of body weight) and Sim (20 mg/kg of body weight) were dissolved in DMSO (final concentration ≤ 5%). Mice from the control group were given CD diet and injected intraperitoneally with DMSO. Mice from the HFD-induced group were fed HFD diet and injected intraperitoneally with DMSO. Mice from the FC group were fed HFD diet and injected intraperitoneally with FC (10 and 20 mg/kg of body weight). Mice from the positive control group were fed HFD diet and injected intraperitoneally with Sim (20 mg/kg of body weight). The intraperitoneal injections were performed three times a week for ten weeks (mouse age from eleven weeks to twenty weeks).
Biochemical parameter analysis
Body weight was recorded twice a week throughout the experiment. All animals were sacrificed by decapitation 6 h after fasting. Blood samples were collected and centrifuged at 4,000 rpm for 15 min at 4℃. Visceral adipocyte tissues (white fat tissues) were dissected immediately, rinsed with cold physiologic saline solution, weighed, frozen in liquid nitrogen and stored at −80℃ for further analysis. Glycerol, TG, and TC were respectively detected via diagnostic assay kits from Nanjing Jiancheng Company following the manufacture's protocol.
Lipid accumulation assay
Differentiated 3T3-L1 adipocytes were treated with FC or Sim for 24 h and fixed with formalin as described [29]. Next, cells were stained with oil red O for 1 h. The cells were treated with isopropanol and lipid accumulation was determined by the absorbance at optical density 490 nm.
Western blot analysis
The cells and visceral adipocyte tissues were lysed with lysis buffer containing a mixture of protease and phosphatase inhibitors in ice condition for 30 min. The protein extracts were harvested by centrifugation at 13,000 rpm for 20 min at 4℃. Protein samples were quantified by using an Enhanced Bicinchoninic Acid Protein Assay Kit (Beyotime, Jiangsu, China). Fifty micrograms of protein was separated on 10% sodium dodecyl sulfatepolyacrylamide gel and electrotransferred to polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% non-fat milk in Tris-buffered saline, and then probed with primary antibodies (1:1,000) for PPAR-α, PPAR-β, PPAR-γ, C/EBP-α, C/EBP-β, SREBP-1c, aP2, LPL, FAS, HSL, AQP-7, ATGL, and β-actin at 4℃ overnight. β-actin was used as a loading control. After three washing with TBST, the membranes were incubated with alkaline phosphatase labeled secondary antibody. Membranes were stained via 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium at room temperature for 10 min. Finally, the relative band intensity was normalized by β-actin and determined by software Quantity One v4.62 (Bio-Rad, Inc., Berkeley, CA, USA).
Quantitative real-time polymerase chain reaction (RT-PCR)
Total RNA was extracted from differentiated 3T3-L1 adipocytes and visceral adipocyte tissue using TRIzol reagent according to the manufacture's protocol (Invitrogen, Carlsbad, CA, USA). First-strand cDNA was synthesized using the cDNA synthesis system (Invitrogen). Quantitative real-time PCR was performed using the Quanti Fast SYBR Green PCR Kit (Qiagen, Valencia, CA, USA). The PCR primers used were shown in Table 1. The threshold cycle (Ct) value for each gene was normalized by β-actin.
Statistical analysis
The data were presented as means ± standard error of the mean of triplicate tests. All results were analyzed via one-way ANOVA followed by the Newman-Keuls test. Calculations were performed using IBM SPSS version 21.0 (IBM Co., Armonk, NY, USA). Differences of p < 0.05 were considered statistical significant.
RESULTS
FC reduces lipid accumulation in differentiated 3T3-L1 adipocytes
As shown in Fig. 1B, the cytotoxic effect analysis indicated that FC at the tested concentrations showed no significant cytotoxicity in 3T3-L1 adipocytes. To evaluate whether FC had potential anti-adipogenic effect, we determined lipid accumulation by the absorbance method in differentiated 3T3-L1 adipocytes. As shown in Fig. 1C, FC could dose-dependently reduce lipid accumulation, and the maximum inhibition was at concentration of 10 µM (p < 0.01). Effect of FC (10 µM) could be comparable to positive reference Sim (10 µM). Glycerol was detected via diagnostic assay kit. As shown in Fig. 1D, FC notably increased the levels of glycerol in the culture supernatants. Effect of FC (10 µM) could be comparable to positive reference Sim (10 µM). These results indicated that FC showed no significant cytotoxicity and reduced lipid accumulation in differentiated 3T3-L1 adipocytes.
FC differentially inhibits the expressions of adipogenesis-related genes and proteins
As shown in Fig. 2A, B, the tested concentrations of FC dose-dependently inhibited the protein expressions of PPAR-γ, but not of PPAR-α and PPAR-β. However, FC significantly suppressed the protein expressions of C/EBP-α, C/EBP-β, and SREBP-1c in differentiated 3T3-L1 adipocytes. The maximum inhibition was at concentration of 10 µM (p < 0.01). Effect of FC (10 µM) could be comparable to positive reference Sim (10 µM). Furthermore, as shown in Fig. 2C, FC treatment markedly down-regulated the mRNA expressions of PPAR-γ, C/EBP-α, C/EBP-β, and SREBP-1c, which showed a similar trend to the changes of the protein expression. Together these results suggested that FC differentially inhibited the expressions of adipogenesis-related factors in differentiated 3T3-L1 adipocytes.
FC suppresses the expressions of lipid synthesis-related genes and proteins
As shown in Fig. 3A, B, the tested concentrations of FC dose-dependently suppressed the protein expressions of aP2, LPL, and FAS in differentiated 3T3-L1 adipocytes, and the maximum inhibition was at concentration of 10 µM (p < 0.01). Effect of FC (10 µM) could be comparable to positive reference Sim (10 µM). Furthermore, as shown in Fig. 3C, FC treatment markedly down-regulated the mRNA expressions of aP2, LPL, and FAS, which showed a similar trend to the changes of the protein expression. These data demonstrated that FC suppressed the expressions of lipid synthesis-related factors in differentiated 3T3-L1 adipocytes.
FC increases the expressions of lipolysis-related genes and proteins
As shown in Fig. 4A, B, the tested concentrations of FC dose-dependently increased the protein expressions of HSL, AQP-7, and ATGL in differentiated 3T3-L1 adipocytes, and the maximum inhibition was at concentration of 10 µM (p < 0.01). Effect of FC (10 µM) could be comparable to positive reference Sim (10 µM). Furthermore, as shown in Fig. 4C, FC treatment markedly up-regulated the mRNA expressions of HSL, AQP-7, and ATGL, which showed a similar trend to the changes of the protein expression. These results demonstrated that FC increased the expressions of lipolysis-related factors in differentiated 3T3-L1 adipocytes.
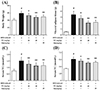
FC decreases both the body weight and visceral adipose tissue weight in HFD-induced obese mice
Five-week-old male C57BL/6 mice were fed HFD diet, the lipid profiles including the body weight (Fig. 5A), visceral adipose tissue weight (Fig. 5B), TG (Fig. 5C), and TC (Fig. 5D) were significantly (p < 0.01) elevated. Then, these obese mice were treated with FC by intraperitoneal injection. As shown in Fig. 5A, B, FC administration markedly decreased both the body weight and visceral adipose tissue weight of HFD-induced obese mice. Meanwhile, the plasma levels of TG and TC were also markedly decreased in the FC group compared with the HFD-induced group (Fig. 5C, D). Effect of FC (20 mg/kg of body weight) could be comparable to positive reference Sim (20 mg/kg of body weight). These data indicated that FC improved lipid accumulation in HFD-induced obese mice.
FC differentially regulates the expressions of lipid metabolism-related factors in HFD-induced obese mice
As expected (Fig. 6A), the data of real-time PCR analysis showed that FC notably down-regulated the mRNA expressions of PPAR-γ, C/EBP-α, C/EBP-β, SREBP-1c, aP2, LPL, and FAS, and also markedly up-regulated the mRNA expressions of HSL, AQP-7, and ATGL in mouse visceral adipocyte tissue, compared with the HFD-induced group. Intriguingly, FC could not regulate the mRNA expressions of PPAR-α and PPAR-β in mouse visceral adipocyte tissue, which showed a similar trend to the changes of the mRNA expression in differentiated 3T3-L1 adipocytes. Furthermore, as shown in Fig. 6B, C, FC administration differentially affected these protein expressions, which showed a similar trend to the changes of the mRNA expression. Together these results demonstrated that FC could dose-dependently and differentially regulate the expressions of lipid metabolism-related factors in HFD-induced obese mice.
DISCUSSION
Obesity is a major risk of serious diet-related chronic diseases, such as hyperlipidemia, hypertension, type II diabetes, atherosclerosis, cardiovascular disease, non-alcoholic fatty liver disease, and metabolic syndrome [123]. In the present study, we indicated that FC could dose-dependently and differentially regulate the expressions of lipid metabolism-related factors in vitro and in vivo. Our results demonstrated that FC had anti-obesity effects via modulating adipogenesis and lipolysis.
Obesity-related syndromes are caused by TG overload and fat deposition [45]. When TG levels are excessive, TG overload will cause lipid accumulation and interfere with the normal function of adipocytes. Excess fat deposition is causally linked to metabolic syndrome abnormalities [67]. On one hand, FC could dose-dependently reduce lipid accumulation in differentiated 3T3-L1 adipocytes. On the other hand, FC could notably increase the levels of glycerol in the culture supernatants. These findings suggested that FC exhibited anti-obesity effects via suppressing lipid accumulation.
Adipogenesis is considered as the complex processes that preadipocytes changed into differentiated mature adipocytes [30]. Pre-adipocytes require the sequential action of adipogenesis-related genes to induce their differentiation into mature adipocytes [29]. The PPARs affect adipocytes differentiation and regulate adipogenesis. PPAR-α and PPAR-β affect the early stage of adipocytes differentiation, while PPAR-γ is involved in the later stage [8]. In the present study, the tested concentrations of FC dose-dependently inhibited PPAR-γ gene and protein productions in differentiated 3T3-L1 adipocytes, but not of PPAR-α and PPAR-β. Furthermore, FC could not regulate the mRNA expressions of PPAR-α and PPAR-β in mouse visceral adipocyte tissue, which showed a similar trend to the changes of the mRNA expression in differentiated 3T3-L1 adipocytes. It is possible that the mechanism of anti-adipogenic effect of FC is distinct from statins. FC could not hydrolyze to the β-hydroxy acid form [3132333435]. Therefore, we evaluated the other transcription factors in the early stage of adipocytes differentiation. C/EBPs and SREBPs might be related to the early stage of adipocytes differentiation process [9]. FC treatment markedly decreased the levels of C/EBP-α, C/EBP-β, and SREBP-1c both in vitro and in vivo. These data demonstrated that FC differentially inhibited the expressions of adipogenesis-related factors.
Emerging studies have suggested that major lipid synthesis enzymes were aP2, LPL, FAS, and so on [101112]. aP2 regulates lipid and glucose metabolism in mature adipose tissue [10]. LPL hydrolyzes lipoproteins from chylomicrons to free TG in serum [11]. FAS catalyzes and synthesizes long-chain fatty acids [12]. FC treatment markedly decreased the levels of aP2, LPL, and FAS both in vitro and in vivo, compared with the other lipid metabolism-related genes. These results indicated that FC suppressed the expressions of lipid synthesis-related factors.
Lipolysis, a process of hydrolysis of TG and TC ester, is mainly measured by HSL, ATGL, AQP-7 and so on [13]. HSL and ATGL break down TG to reduce the visceral adipose tissue weight [14]. AQP-7 is an adipocyte membrane channel protein, and it affects the release of glycerol from adipocytes into serum [15]. In the present study, FC administration markedly decreased both the body weight and visceral adipose tissue weight of HFD-induced obese mice. Meanwhile, the levels of TG and TC were also markedly decreased. FC also notably increased the levels of HSL, AQP-7, and ATGL both in vitro and in vivo. These data demonstrated that FC could decrease TG and reduce the weight of HFD-induced obese mice.
In summary, the present study elucidated that FC exhibited anti-obesity effects via modulating adipogenesis and lipolysis in differentiated adipocytes and HFD-induced obese mice. The mechanisms were involved in regulating the expressions of lipid metabolism-related factors, attenuating adipogenesis, and reducing lipid accumulation.






 XML Download
XML Download