

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Testes, as a critical component of the male reproductive and endocrine tract, the primary functions is to produce sperm and androgens. Mammalian testes consists of several well-defined cell types, including germ cells of different stages, Sertoli cells and Leydig cells. Each of them is configured with a different location and has specific functions: Germ cells produce sperm and are located in the seminiferous tubules; Leydig cells produce sex hormones and are distributed in the connective tissue of the convoluted seminiferous tubules; Sertoli cells form the basement membrane of the seminiferous tubules and offer the environment necessary for the differentiation and maturation of germ cells [1]. The importance of Leydig cells in male reproduction is exemplified by the fact that loss of function of gene such as the luteinizing hormone receptor (LHR) [2], insulin-like factor 3 (Insl3) [3], and enzymes involved in testosterone biosynthesis [4].
C-type natriuretic peptide (CNP) and its receptor (natriuretic peptide receptor 2, NPR2) play a role as an oocyte maturation inhibitor in mammalian [5]. Recent studies suggested that CNP can improve oocyte maturation and developmental competence in vitro [6]. Compared with numerous studies about CNP/NPR2 on female reproduction, there is little research about CNP/NPR2 in male reproduction. Previous studies reported that CNP regulates blood-testis barrier dynamics and is related with spermatogenesis [7]. Sogawa et al [8] investigated phenotype of NPR2-defcient short-limbed-dwarfs mice, and found that the developmental onset and acquisition of spermatogenic function is delayed in NPR2 mutant mice. Subsequently, two studies reported that CNP/NPR2 is related with sperm motility, acrosome reaction and induces sperm attraction for fertilization, thus regulating the reproductive function of males [910].
However, the function of CNP/NPR2 in male reproduction remains largely unknown. In this study, we aimed to study the role of CNP/NPR2 on cell proliferation, testosterone secretion and related regulatory mechanisms in mouse Leydig cells.
MATERIALS AND METHODS
1. Chemicals and mice
All chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated. Male Kunming mice were purchased from the Laboratory Animal Central of Jiujiang University. All mice were fed a typical diet of lab chow and housed in a single room under conditions of constant temperature (25℃– 28℃), humidity (55%±5%) and lighting (12 hours light, 12 hours dark cycle) [11]. All procedures were approved by the Committee for the Ethics on Animal Care and Experiments of Jiujiang University (approval No. SYXK(GAN)2017-0001).
2. Isolation and cultures of Leydig cells
Leydig cells were isolated from the testes of 42- to 49-day-old Kunming mice and cultured as previous report [12]. The purity of Leydig cells was assessed by 3β-hydroxysteroid dehydrogenase (3β-HSD) staining using the modified Wiebe method. Simply, Leydig cells were fixed in 1% paraformaldehyde for 20 minutes and then washed with phosphate buffer saline (PBS) for three times. After washed the cells were incubated with 1 mL PBS containing 1 mg bovine serum albumin, 1.5 mg nicotinamide adenine dinucleotide, 0.2 mg nitroblue tetrazolium and 0.25 mg dehydroepiandrosterone for 2 hours at 37℃ in the dark. Then the cells were gently rinsed with PBS and observed microscopically. During all the culture, the cells were cultured with Dulbecco's modified Eagle's medium/F-12 containing 10% fetal bovine serum with or without CNP (CNP were used to active CNP/NPR2 signaling pathway), unless otherwise stated.
3. Transfection of cells with shRNA-natriuretic peptide receptor 2 lentiviral
shRNA-NPR2 and shRNA-negative lentiviral vector were purchased from Genechem (Shanghai, China). The mouse Leydig cells were seeded into 6-well plates, which were cultured to 30% to 40% confluence and infected by addition of 1×108 TU/mL lentivirus, 5 µg/mL polybrene and complete medium. After 12 hours, the lentivirus solution was replaced by complete culture medium and cultured for 36 hours. And the cells were observed under a fluorescence microscope to evaluate the transfection efficiency. The efficiency and specificity of siRNA mediated knockdown were examined by Western blot and quantitative reverse transcription polymerase chain reaction (RT-qPCR). After determine the NPR2 knockdown efficiency, the cells were cultured for subsequent experiments.
4. Cell proliferation assay
After culturing with CNP for 24 hours, Cell Counting Kit-8 (CCK8) was added to the cells (10 µL/well). Then the cells were incubated for 1 hour at 37℃ and measured at 450 nm by a Microplate Reader (Bio-Rad 680, Hercules, CA, USA). The experiments were performed in triplicate.
5. Cyclic guanosine monophosphate measurement
After 30 minutes of treatment, the cells were collected to measure the cyclic guanosine monophosphate (cGMP). The cGMP measurement was according to the procedure described as our previous report [6]. The levels of cGMP were determined using cGMP-EIA kits obtained from Cayman Chemicals (Ann Arbor, MI, USA). Each sample was measured in triplicate.
6. Testosterone measurement
After 24 hours of treatment, mouse Leydig cells were counted. The concentration of testosterone in the culture supernatants (100,000 cell/mL culture supernatant) was measured with ELISA kits (Ji Yin Mei; Co. Ltd., Wuhan, China) according to the manufacturer's instructions. Each sample was measured in triplicate.
7. Cell cycle analysis
Mouse Leydig cells of the respective experimental groups were collected and fixed in ice-cold 70% ethanol overnight at 4℃ after the cells were treatment and count. Then, the cells were washed with PBS and stained with propidium iodide (PI) solution for 20 minutes at 25℃ in the dark. Finally, the cells were analyzed by flow cytometry using a BD FACS Calibur system and Mod Fit LT for MacV3.0 software. For each analysis, a minimum of 10,000 cells were analyzed. Each sample was measured in triplicate.
8. Cell apoptosis assay
The apoptotic rate was determined using the phycoerythrin (PE) and PI double-staining apoptosis analysis kit (Nanjing Key Gen Biotech, Nanjing, China). After washes by PBS for two times, followed by centrifugation, the cells were suspended in 500 µL binding buffer. Then, 5 µL Annexin PE and PI staining solution were added and incubated at 25℃ for 10 minutes. The mixture was then analyzed by flow cytometry within 60 minutes. Each sample was measured in triplicate.
9. RNA isolation and quantitative reverse transcription polymerase chain reaction
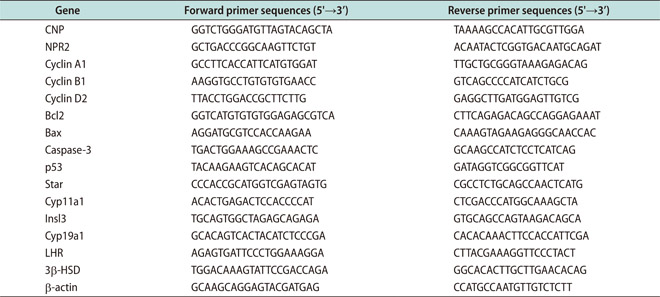
Total RNA was extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA). First-strand cDNA was synthesized according to the manufacturer's instructions (PrimeScript® RT reagent Kit). RT-qPCR was performed using an ABI StepOnePlus Real-time Detection System (ABI, Foster, CA, USA) and SYBR Green qPCR SuperMix (Invitrogen). Each experiment was repeated independently at least three times, and the fold change in the expression of each gene was analyzed using the 2-ΔΔCT method. All the primers were used as Appendix and the β-actin was used as internal control.
10. Immunohistochemistry
Testes of 42- to 49-day-old Kunming mice were fixed in 4% paraformaldehyde in PBS for one week, dehydrated through a graded ethanol series, and embedded in paraffin. Sections 7 mm thick were mounted onto glass slides precoated with Poly-L-Lysine Solution and incubated overnight at 37℃. After dehydrating, samples were placed in citrate buffer (pH=6.0). Antigen retrieval was performed by treating samples in a microwave oven at 92℃ for 15 minutes; slides were cooled and then washed in PBS. The sections were pretreated with 3% (vol/vol) H2O2 in methanol to quench endogenous peroxidase activity. After being washed with PBS, sections were incubated with 10% goat serum for 30 minutes at 37℃. After blocking, sections were incubated overnight at 4℃ with rabbit polyclonal antibody against NPR2 (Sigma-Aldrich; 1:50 dilutions) in a humidified chamber. After washing, followed by incubation with biotinylated anti-rabbit immunoglobulin G antibody (Beijing 4A Biotech Co., Ltd, Beijing, China) at 37℃ for 1 hour, and then sections were incubated with horseradish peroxidase-labeled streptavidin at 37℃ for 30 minutes. Thereafter, positive reactions were visualized with a diaminobenzidine-peroxidase substrate and 30 seconds counterstaining with hematoxylin. Finally, the sections were counterstained with dehydrated and mounted. Slides were imaged using a digital microscope (BA400; Motic, Wetzlar, Germany).
11. Western blot analysis
Western blot analysis The extraction of cell proteins was used radioimmunoprecipitation assay buffer. Protein determination was performed by the bicinchoninic acid (BCA) assay. 200 µg total protein per sample were separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and electro transferred to a to polyvinylidene difluoride membrane. After incubation in blocking buffer for 1 hour at room temperature, the membrane was incubated overnight at 4℃ with the primary antibodies (p-AKT, AKT, β-actin, 1:1,000, Cell Signaling Technology, Beverly, MA, USA; NPR2, 1:500, Sigma-Aldrich; Cyp11a1, Star, Bax, Bcl-2, 1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA). After washing, the membranes were incubated with a secondary antibody conjugated to horseradish peroxidase at 25℃ for 1 hour. Finally, immunoreactive bands were visualized using a Super Signal West Pico kit according to the IL, USA) instructions. The β-actin was used as internal control and the protein band densities were semiquantified by densitometric analysis using ImageJ (ver. 1.49).
12. Caspase-3 activity measurement
Caspase-3 activity was measured using a caspase-3 Activity Colorimetric Assay Kit. Simply, after treatment, the cells were harvested by centrifugation and incubated in lysis buffer on ice for 15 minutes. The lysate was then centrifuged at 15,000 rpm and 4℃ for 15 minutes, and the protein content was determined using the BCA Protein Assay Kit according to the manufacturer's (Nanjing Biobox Biotech, Naniing, China) instructions. Then, 100 mg of protein in each sample was incubated with the caspase-3 substrate (200 mM final concentration) at 37℃ in a microplate for 4 hours. The samples were measured at 405 nm using a microplate reader (Bio-Rad 680).
13. Statistical analyses
All experiments were replicated at least three times for each group, and the data are presented as the mean±standard error of mean. The data were analyzed by ANOVA, followed by Fisher's least significant different test and independent samples Student t-test, with the SPSS software ver. 13.0 (SPSS Inc., Chicago, IL, USA). Differences were considered significant at p<0.05.
RESULTS
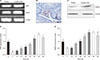
1. The expression pattern of C-type natriuretic peptide/natriuretic peptide receptor 2 at different postnatal stages of testes
Previous studies reported that CNP is mainly located and expressed in rat Leydig cells [1314]. But the expression and location of NPR2 in rodent are still unknown. Here, we first detected the location and expression of NPR2 in adult mouse testes by semiquantitative RT-PCR, Western blot and immunohistochemistry. Results showed that NPR2 is mainly expressed in Leydig cells (Fig. 1A–1C). Moreover, CNP is also mainly expressed in Leydig cells (Fig. 1A). In addition, we detected the expression pattern of CNP and NPR2 at different time of testes after born (0, 7, 14, 21, 28, 35, 42, 49, and 56 days) by RT-qPCR. Result showed that CNP and NPR2 have a similar expression patterns in the mouse testis at different postnatal stages (Fig. 1D, 1E). The mRNA expression of CNP and NPR2 has a higher expression levels on 0 day, reach its lowest level on postnatal 7 days, and then increases again from 14 to 56 days (Fig. 1D, 1E).
2. Effects of C-type natriuretic peptide on cyclic guanosine monophosphate production, natriuretic peptide receptor 2 expression, cell proliferation, testosterone secretion in mouse Leydig cells
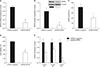
To measure the effects of CNP on cGMP production, NPR2 expression, cell proliferation, testosterone secretion in mouse Leydig cells, the cells were treated with different concentrations of CNP (0, 15, 30, 60, 120 nM) to measure cGMP production, detect NPR2 expression, cell proliferation and testosterone secretion. The results showed that CNP enhances cGMP production in a dose-dependent manner, and the lowest effective concentration is 15 nM (Fig. 2A). Moreover, RT-qPCR results showed that CNP also increases NPR2 expression in dose-dependent manner and the lowest effective concentration is 30 nM (Fig. 2B). Meanwhile, CCK8 results showed that CNP enhances cell proliferation in a dose-dependent manner, and the lowest effective concentration is 30 nM (Fig. 2C). In addition, ELISA results showed that CNP promotes the testosterone secretion and the lowest effective concentration is 30 nM (Fig. 2D).
3. Natriuretic peptide receptor 2 knockdown inhibits cyclic guanosine monophosphate production and cell proliferation in mouse Leydig cells
To investigate role NPR2 on cell proliferation, the mouse Leydig cells were transfected with shRNA-NPR2 and shRNA-negative lentivirus. And the NPR2 knockdown or shRNA-negative mouse Leydig cells were treated with 30 nM CNP for 24 hours. RT-qPCR and western blot showed that shRNA-NPR2 lentiviral effectively suppresses NPR2 expression in both mRNA and protein levels (Fig. 3A, 3B). Moreover, ELISA showed that NPR2 depleted Leydig cells have a lower expression levels of cGMP (Fig. 3C). CCK8 results indicated that a decreased cell viability is observed in NPR2 knockdown group compared with the control group (Fig. 3D). Flow cytometry analysis showed that the S phase of cells in the NPR2 knockdown group is higher compared the control group (Table 1). In addition, the mRNA expression of cell cycle factors (cyclin A1, cyclin B1, and cyclin D2) is downregulation in NPR2 knockdown mouse Leydig cells compared the control group (Fig. 3E).
4. Natriuretic peptide receptor 2 knockdown increases apoptosis in mouse Leydig cells
To determine the effect of NPR2 knockdown on mouse Leydig cells apoptosis, the cells were cultured in culture medium with 30 nM CNP for 24 hours and detected by flow cytometry, RT-qPCR and Western blot. The results showed that the apoptosis rate of the shRNA-NPR2 group is significantly higher than shRNA-negative group (Table 2). Furthermore, the mRNA levels of apoptosis-related genes (p53, caspase-3, Bcl-2, Bax) were quantitatively analyzed. The mRNA levels of p53 and caspase-3 and Bax are obviously upregulated. However, the Bcl-2 expression is not altered significantly (Fig. 4A). Moreover, NPR2 knockdown increases the caspase-3 activity and the BAX protein expression, but not Bcl-2 (Fig. 4C, 4D). In addition, we also detected the phosphorylation of AKT, which is important for cell proliferation. The results showed that the pAKT of shRNA-NPR2 group is significantly decreased compared with shRNA-negative group (Fig. 4B).
5. Natriuretic peptide receptor 2 knockdown decreased testosterone secretion in mouse Leydig cells
To assess the role of NPR2 on steroid hormone levels, we measured the concentrations of testosterone in culture medium with 30 nM CNP or 2 IU human chorionic gonadotropin (hCG) after cultured for 24 hours. The results showed that both CNP and hCG can promote testosterone secretion and the expression of steroidogenic enzymes (Fig. 5B–5D). Moreover, hCG seems to enhance the CNP/NPR2 expression (Fig. 5A). After transfection, the levels of testosterone are significantly lower in the shRNA-NPR2 group than negative group both in CNP and hCG treatment (Fig. 5B). We further analyzed the mRNA expression of several genes encoding steroidogenic enzymes, including Star, Cyp11a1, 3β-HSD, Insl3, and LHR. The result showed that the mRNA levels of Star, Cyp11a1, Insl3, and 3β-HSD significantly decreased, and the protein levels of Star and Cyp11a1 also induced by CNP and hCG, whereas LHR are no obvious change in mRNA level (Fig. 5C, 5D).
6. The mechanism of C-type natriuretic peptide, testosterone and estradiol in natriuretic peptide receptor 2 expression in mouse Leydig cells
Because testosterone promotes NPR2 expression directly via androgen receptor (AR) or indirectly via estrogen receptor (ER) by promoting estrogen production [15], and Leydig cells can convert a bit of testosterone to estrogen. We hypothesized that CNP promotes NPR2 expression via AR or ER in mouse Leydig cells. To test this hypothesis, the mouse Leydig cells were treated with 30 nM CNP, 30 nM testosterone, or 30 nM estrogen in the presence or absence of 500 nM AR inhibitor flutamide and 500 nM ER inhibitor ICI182780 for 24 hours during the culture.
Results showed that flutamide partly inhibits the CNP- and testosterone-induced NPR2 expression in mouse Leydig cells, but no obvious effects on estrogen-induced NPR2 expression (Fig. 6A). At the same time, ICI182780 successfully inhibits the expression of NPR2 induced by estrogen, but no obvious changes is observed in other groups (Fig. 6A). We also detected the CYP19a1 (the enzyme responsible for androgen aromatization to estrogen) expression by RT-qPCR. The results showed that all the treatment have no obvious effects on CYP19a1 expression (Fig. 6B).
DISCUSSION
Numerous studies have demonstrated that CNP/NPR2 plays an important role in female reproduction. However, the functions of CNP/NPR2 in male reproduction, especially in Leydig cells, remain poorly understood. In this research, we aimed to study the role of CNP/NPR2 on cGMP production, cell cycle, apoptosis, testosterone secretion and related regulatory mechanism in mouse Leydig cells.
We first detected NPR2 location in male mouse testis and found NPR2 is mainly expressed in Leydig cells. As far as know, this is the first report that NPR2 is mainly expressed in mouse Leydig cells. We also found that CNP was expressed in mouse Leydig cells, this is similar with the previous studies in rat [1314]. We also detected CNP/NPR2 expression from born to sexual maturity and showed age-related increase, although the expression level of CNP/NPR2 is higher at birth. Higher expression levels of CNP/NPR2 at birth maybe that:1) CNP/NPR2 may play an important role in fetal development, especially the rapid growth of the fetus (CNP/NPR2 is a key regulatory factor for bone growth) [1617]; 2) Higher expression levels of CNP/NPR2 on 0 day indicated that they may have an intimate relation ship with the onset of spermatogenesion [8]. This results are further supported by a previous study in rat [7].
The change in CNP/NPR2 expression may imply that CNP/NPR2 may be involved in the testicular development, testosterone secretion, spermatogenesis in the male reproduction as previous reports [781318]. As NPR2 is expressed in Leydig cells of male mice testes and the binding CNP and NPR2 generates cGMP to regulate cell function. So, we used Leydig cells as cell model and added CNP in the culture medium to active CNP/NPR2 signaling pathway during the following research. We found that CNP promotes cGMP production in Leydig cells, suggesting that NPR2 is successfully active by CNP.
The effect of CNP/NPR2 signaling pathway on Leydig cell proliferation and cycle have never reported. Our study revealed that CNP enhances cell proliferation in a dose-dependent manner. NPR2 knockdown significantly reduces the proliferation activity and the cells are arrested at the S phase. This may be because CNP/NPR2 is related with expression of cell cycle factors (cyclin A1, cyclin B1, and cyclin D2). Cyclin A1 and cyclin D2 are key regulators associated with the S-to-G2/M transition. Moreover, cyclin B1 is an important gene related to regulating the normal cell-cycle progression and plays a vital role in mitotic exit and the beginning of the subsequent cell division. Thus, the cell-cycle results imply that CNP/NPR2 is involved in the growth and proliferation in Leydig cells. In addition, some studies report that CNP is a growth inhibitor in smooth muscle cells [1920], and some studies reported that CNP promotes the growth of the bone [17]. This difference may be related to cell types or the indirect effects of CNP, such as testosterone synthesis [21] and insulin-like growth factor I production [16].
Previous study showed that NPR2 gene knockout mice have smaller size [16] and smaller testis [8] compared with the normal mice. However, whether CNP/NPR2 regulates mouse Leydig cells viability and apoptosis remains unclear. In the current study, we found that CNP effectively promotes the viability of mouse Leydig cells. At the same time, NPR2 depletion remarkably induces apoptosis in mouse Leydig cells. To clarify this result further, we measured the expression of key molecules associated with apoptosis via RT-qPCR and Western blot. As an important transcription factor, p53 can promote cell apoptosis, and elevated p53 mRNA levels result in higher apoptosis rates in Leydig cells [22]. Caspase-3, acts as an executioner in caspase-mediated apoptosis, is extensively involved various cell apoptosis. Moreover, in the Bcl-2 family, Bcl-2 plays an anti-apoptotic role, whereas Bax has a proapoptotic effect [2324]. In the present study, the expression of Bax, p53 and caspase-3 was significantly upregulated after NPR2 disruption, whereas the expression Bax levels appeared unchanged. Based on these results, we concluded that the apoptosis cells in mouse Leydig cells caused by disrupting NPR2 is closely correlated with these molecules. In addition, PI3K-AKT pathway plays an key role in cell proliferation and survival [25] and NPR2 knockdown inhibits the activation of AKT in in mouse Leydig cells. This also showed that NPR2 regulates cells viability and apoptosis in another perspective.
Given the importance of Leydig cells in secretion of testosterone, we evaluated the CNP/NPR2 signaling pathway on testosterone synthesis in mouse Leydig cells. Our results indicated that CNP promotes testosterone secretion in mouse Leydig cells as a previous report [21] and NPR2 silencing inhibits testosterone production. The possible reason for the reductions of testosterone could be the decreases in the Star, Cyp11a1 and 3β-HSD, which is essential for testosterone biosynthesis [426]. Moreover, the downregulation of cGMP may also inhibits testosterone production [27]. However, the expression levels of LHR was no obvious after NPR2 knockdown in Leydig cells. In addition, we also found that activated LH signal by hCG induces the CNP/NPR2 expression and NPR2 disruption partly reduces the hCG-induced testosterone secretion. This result suggested that CNP/NPR2 signal maybe related with LHR signal during the testosterone secretion. Thus, we inferred that CNP/NPR2 may play an important role in the development of male reproductive tissues as well as promoting secondary sexual characteristics, such as increased muscle and bone mass by promoting testosterone production.
We also found that CNP promotes NPR2 expression in mouse Leydig cells. And previous studies report that testosterone enhances NPR2 expression in cumulus cell [1528]. So, we hypothesized that CNP-induced NPR2 expression is by promoting testosterone production, which further enhances NPR2 expression via AR in mouse Leydig cells. Consequently, we inhibited AR by flutamide during the culture with CNP and testosterone. The result that inhibited AR by flutamide partially improves NPR2 expression in Leydig cells. We therefore speculated that testosterone is an important factor in regulating CNP/NPR2 in Leydig cells as previous report in ovary [15]. As Leydig cells can convert a bit of testosterone to estrogen [29] and estrogen promotes NPR2 expression via ER [2830], we detected the role of ER on CNP- or testosterone-induced NPR2 expression. We found inhibited ER by ICI182780 has no obvious effects on NPR2 expression in mouse Leydig cells and this result is further supported by the result of CYP19a1 mRNA expression. These results indicated that CNP promotes testosterone secretion, which further induces NPR2 expression via AR in mouse Leydig cells.
CONCLUSIONS
Our study highlights the important role of CNP/NPR2 in mouse testes. We found that CNP/NPR2 is related with cell proliferation and testosterone secretion in mouse Leydig cells. In addition, CNP can promote NPR2 expression via AR in mouse Leydig cells. Our findings not only provided new insights into the function of CNP/NPR2 in female reproduction but also provide potential therapeutic targets for treating reduced testosterone.



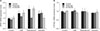


 XML Download
XML Download