

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Semen analysis is a first level diagnostic investigation for male infertility (as well as for the testicular function), since it is able to give indication on sperm fertility potential. However, the implementation of several assisted reproductive techniques (ARTs) has led to the observation that normal sperm parameters are not always capable of inducing pregnancy in a healthy woman. For this reason, several studies on sperm functional capability and bio-functional sperm parameters have been developed [12]. Nonetheless the evaluation of these variables is often insufficient to identify causes of male infertility [3]. To overcome this pitfall, molecular biologists have recently tried to study the epigenetic origin of male infertility and embryonic anomalies [4].
Epigenetics study meiotic and/or mitotic inherited changes affecting gene expression without a DNA sequence modification [5]. These studies allow in some cases to better evaluate cases of so-called idiopathic infertility. DNA methylation, chromatin remodeling and residual histone modifications represent the major epigenetic changes occurring at the sperm level [67]. Recently, sperm RNAs seem also to play an important role [8].
This article reviews the implications of epigenetic modifications on male fertility, fertilization, embryonic development and ART outcomes, highlighting the possible mechanisms involved. To accomplish this, we performed a search on Pubmed, Science-direct, Ovid and Scopus, using the following key words: sperm DNA methylation, sperm DNA methylation and assisted reproduction technology, sperm chromatin remodeling, sperm chromatin remodeling and assisted reproduction technology, sperm histone modification, sperm histone modification sperm RNAs and assisted reproduction technology.
Go to : 
DNA METHYLATION
DNA methylation is a dynamic mechanism of gene expression regulation that occurs on dinucleotides formed by cytosine and guanine. These dinucleotides are grouped in the so called differently methylated regions (DMRs) that are often placed near gene regulatory regions, such as the promoter [9]. The methylation processes are regulated by DNA methyltransferases [9]. Hypermethylation suppresses gene expression, since methyl groups prevent the recruitment of transcription factors and DNA polymerases. Conversely, hypomethylation promotes gene expression [710].
DNA methylation includes global DNA methylation, methylation of imprinted and non-imprinted genes and of methylation of repetitive elements.
A global DNA hypomethylation seems to occur in association with ART [11]. Furthermore, an increased risk of imprinted disorders has been reported after these procedures, thus leading to the hypothesis that ART itself may cause a loss of methylation [11]. Some studies described a significantly different global DNA methylation in sperm from patients with oligoasthenoteratozoospermia (OAT) compared to controls [12], indicating that such epigenetic abnormality may reflect on human fertility. It cannot be excluded whether a change in sperm global DNA methylation may primary affect ART outcome. Indeed, sperm global DNA methylation has been related with chromatin injury in samples from OAT patients [1213] and, interestingly, chromatin integrity seems to impact on ART (see section “CHROMATIN REMODELING”). Thus, sperm global methylation may indirectly influence ART outcome.
Methylation of repetitive elements (e.g., ALU and LINE1) at the sperm level has been reported to influence ART outcome. More in detail, ALU methylation impacts on pregnancy and live-birth rates in couples with male-factor and combined infertility, being significantly lower in sperm samples that led to abortion compared to those leading to pregnancy and live-birth [14].
Imprinted genes show parent-specific activity (they are functionally haploids), which makes them particularly vulnerable to epigenetic dysregulation. In greater detail, both paternal and maternal alleles undergo to demethylation after fertilization. Then, genetic reprogramming consisting in new specific methylations takes place in the embryo [15]. Imprinted genes escape epigenetic reprogramming after fertilization, thus allowing the possible transmission of aberrant methylation patterns into the offspring.
Imprinting has been extensively studied in humans and mice. The role of imprinted genes in processes involved in embryo and placenta development and growth is well recognized in mice (Table 1) [16]. Since humans and mice have common placental physiology [17], these mechanisms may occur also in humans. Knowledge in humans mainly derives from pediatric growth disorders. However, epigenetic studies in human sperm and placenta allow to understand the role of imprinted genes in human fertilization. Accordingly, impaired methylation of imprinted genes has been reported in infertile patients [18]. Furthermore, modifications in sperm DNA methylation of such genes play a role in the development of male infertility and negatively impact on the ART outcomes, leading to abortion [19]. Meta-analytic data provide evidence for lower H19 and higher MEST and SNRPN sperm methylation in infertile patients compared to fertile controls [18].

Table 1
Imprinted genes involved in placenta and embryo development and growth in mice [16]
| Process | Imprinted gene |
|---|---|
| Placenta establishing | Peg10 |
| Nutrient transport capacity and surface area for exchange | Igf2, Grb10 |
| Fetal growth | Igf2, Igf2r, Cdkn1c, Grb10 |
![]()
A list of imprinted genes involved in sperm quality is shown in Table 2 [18192021222324252627]. A focus on the role of H19 DMR impaired methylation rate in human fertility is reported below.
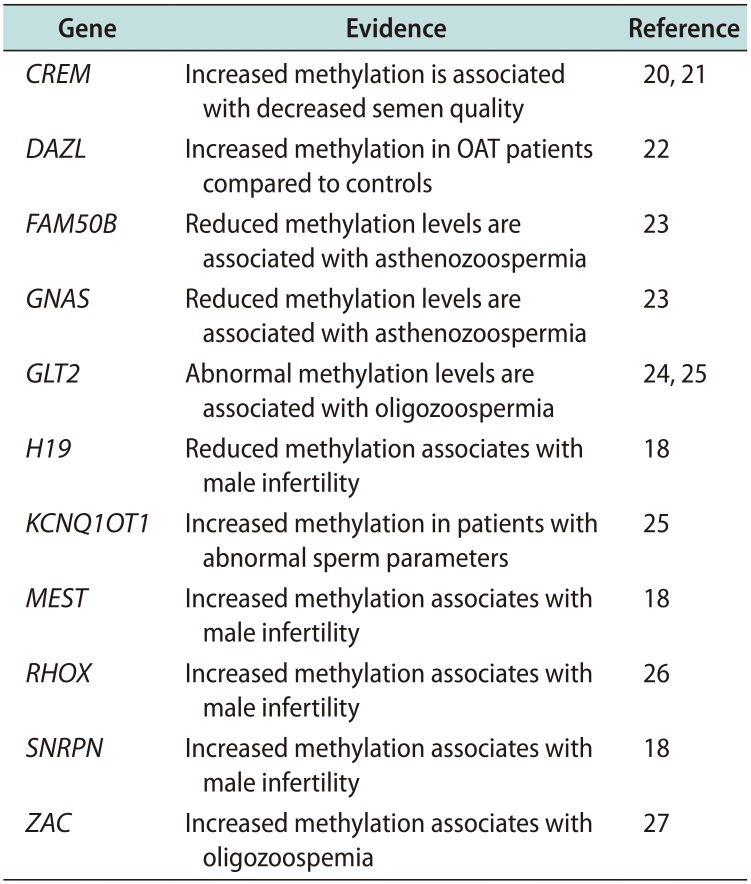
Table 2
Imprinted genes involved in sperm quality
| Gene | Evidence | Reference |
|---|---|---|
| CREM | Increased methylation is associated with decreased semen quality | 2021 |
| DAZL | Increased methylation in OAT patients compared to controls | 22 |
| FAM50B | Reduced methylation levels are associated with asthenozoospermia | 23 |
| GNAS | Reduced methylation levels are associated with asthenozoospermia | 23 |
| GLT2 | Abnormal methylation levels are associated with oligozoospermia | 2425 |
| H19 | Reduced methylation associates with male infertility | 18 |
| KCNQ1OT1 | Increased methylation in patients with abnormal sperm parameters | 25 |
| MEST | Increased methylation associates with male infertility | 18 |
| RHOX | Increased methylation associates with male infertility | 26 |
| SNRPN | Increased methylation associates with male infertility | 18 |
| ZAC | Increased methylation associates with oligozoospemia | 27 |
CREM: cAMP responsive element modulator, DAZL: deleted in azoospermia-like, FAM50B: family with sequence similarity 50, member B, GNAS: guanidine nucleotide-binding protein, alpha-stimulating activity polypeptide 1, GLT2: gene trap locus 2 (also known as MEG3, maternally expressed gene 3), H19: imprinted maternally expressed non-coding transcript, KCNQ1OT1: KCNQ1-overlapping transcript 1, MEST: mesoderm-specific transcript, mouse, homolog of, RHOX: reproductive homeobox X-linked, SNRPN: small nuclear ribonucleoprotein polypeptide N, ZAC: Zac tumor suppression gene (also known as PLAGL1, pleomorphic adenoma gene-like 1).
![]()
1. H19/IGF2
H19/IGF2 represents the first historically characterized couple of imprinted genes [28]. Both H19 and IGF2 map on the 11p15.5 chromosome. H19 encodes for a long non-coding RNA (lncRNA), which negatively modulates human placental trophoblast cell proliferation [29]. In particular, lncRNA H19 targets miR-675, which, in turn, represses IGF1R transcription [30]. IGF2 encodes for a growth factor which is able to activate the insulin-like growth factor-1 receptor (IGF1R) and to promote fetal and placental growth [31323334]. In contrast to IGF1, which is preferentially expressed after birth, IGF2 is mainly produced in the early embryonic development. On these basis, it might be hypothesized that a low sperm IGF2 gene expression might negatively influence the outcome of pregnancy, though no study has investigated this aspect so far.
IGF2 gene expression is regulated by the H19 DMR methylation rate. In the maternal allele, H19 DMR is unmethylated, thus allowing H19 expression and preventing the accession to the IGF2 gene enhancer. In the paternal allele, the H19 DMR methylation leads to IGF2 gene expression [35]. Several studies described the occurrence of low H19 DMR methylation rates in infertile patients [1836373839]. At the sperm level, this may lead to low IGF2 gene expression, thus affecting the outcome of pregnancy.
A study carried on 119 human placentas (56 from in vitro fertilization [IVF], 41 from intracytoplasmic sperm injection [ICSI] and 22 from natural conception) found 1.9 and 1.8-fold higher H19 messenger RNA (mRNA) levels, respectively in ICSI and IVF-derived placentas compared with those coming from natural conception. In addition, IGF2 mRNA levels resulted significantly lower in couples undergoing ART compared to that of controls. Furthermore, although both birth and placental weight were lower in the ART group, they were not significantly different compared to controls [40]. This study provides evidence for abnormal H19 and IGF2 expression in ART placentas. Due to abnormal H19 DMR methylation rate observed in infertile patients [18], men undergoing to ART programs may likely show H19 gene hypomethylation. This might explain the abnormal H19 and IGF2 expression in ART placentas [40].
Finally, IGF1R has been recognized to play a role in human sperm capacitation [41]. Therefore, due to the inhibitory effect that H19 has on IGF1R gene transcription [30], it might be hypothesized that low H19 DMR methylation rates may also impair IGF1R gene expression, thus affecting human sperm capacitation.
Taking all this into account, H19/IGF2 gene expression may influence human fertility (Fig. 1). Other studies relating pregnancy rate with IGF2 and IGF1R mRNA levels in human spermatozoa may further clarify their role.
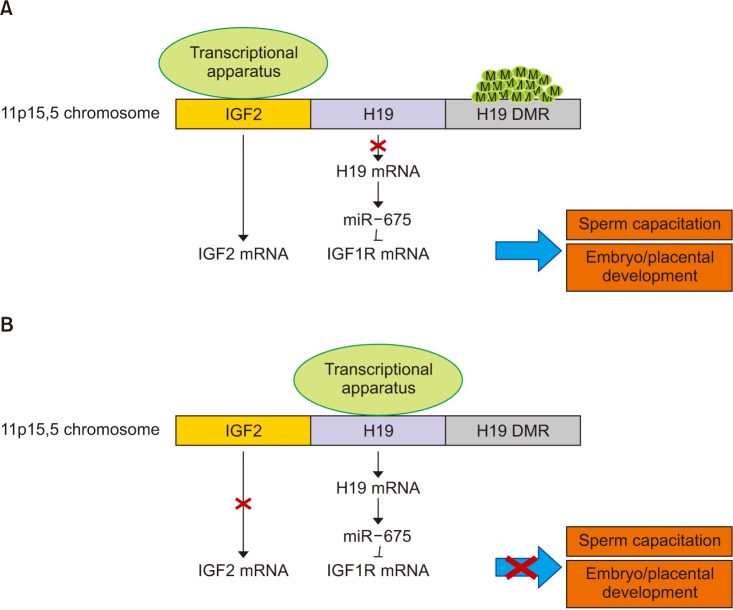 | Fig. 1Possible influence of H19/IGF2 expression on male fertility. (A) In the paternal allele of healthy fertile men, H19 DMR methylation leads to IGF2 expression. The repression of H19 transcription increase IGF1R expression. Both IGF2 and IGF1R transcripts are involved in sperm capacitation, embryo and placental growth. (B) In infertile men, low H19 DMR methylation rates lead to repression of both IGF2 and IGF1R transcription, thus negatively impacting on sperm capacitation, embryo and placental growth. H19: Imprinted maternally expressed non-coding transcript, IGF2: insulin-like growth, DMR: differentially methylated region, IGF1R: insulin-like growth factor 1 receptor, factor 2.
|
Go to :
CHROMATIN REMODELING
Spermatozoa are highly specialized cells. During spermatogenesis, almost all chromatin histones (90%–95%) are replaced by protamines, that are small, arginine-rich, nuclear proteins [42]. This process, typical of spermatozoa, ensures a significant DNA compaction, a decreased susceptibility to external insults and it represents a gene silencing mechanism. During early stages of protamination, there is an increased acetylation of histones that firstly promotes the action of DNA topoisomerase enzyme, followed by the substitution of histones with transition proteins (TP1 and TP2) [43]. These DNA binding proteins facilitate histones removal and subsequent replacement of protamines 1 (P1) and 2 (P2), which are expressed in equal amounts [44]. After sperm chromatin protamination, three levels of DNA organization can be recognized: toroidal structures formed by protamines (90%–95%), nucleosomes, involved in the primary phases of embryonic development (5%–10%) and DNA segments called matrix attachment regions (MARs), in which neither toroidal structures nor nucleosomes are present [43]. MARs provide chromatin with structural support, working as promoter in paternal pronuclear formation after fertilization, as well as contributing to normal embryogenesis.
Several studies reported that an erroneous sperm protamination or an alteration in one of the three DNA organization levels leads to a lower sperm quality and decreased pregnancy rate and embryo quality following ARTs [642]. In greater detail, a P1/P2 ratio ranging from 0.54 to 1.43 has been reported in normozoospermic men [45]. Abnormal low or high P1/P2 ratio is associated with poor sperm [454647] and embryo quality and with low fertilization and pregnancy rates [48]. Indeed, abnormal sperm protamination may result in insufficient sperm DNA condensation, thus making it susceptible to the oxidative damage [49]. Accordingly, impaired sperm protamination has been related to increased DNA fragmentation [4850]. Furthermore, several factors negatively affecting male fertility and sperm quality have been found to impair sperm chromatin compactness, such as overweight and obesity [51], nicotine [5253], male accessory gland infections [54] and varicocele [55]. Lower chromatin compactness and poor sperm quality have been described in patients with low testicular volume [56].
In conclusion, recent scientific evidence showed that anomalies in protamine content may affect epigenetic information transmitted by the paternal DNA [49] and, not surprisingly, sperm protamination has been already suggested as a target to predict ART outcomes [57].
Go to :
HISTONE MODIFICATION
After protamination, the residual histones (5%–10%) are organized into nucleosomes that are octamers composed by two dimers of H3–H4 and two of H2A-H2B histones. These residual histones play a crucial role in sperm epigenetics. In fact, they might undergo chemical modifications that might drastically alter their DNA binding abilities, favoring the action of transcription factors and therefore gene activation or silencing [58]. These chemical modifications are mainly referred to lysine or serine residues located on the histone tail, undergoing to methylation, acetylation, phosphorylation or ubiquitination [42]. Moreover, histones may have variants such as the H2B, that is a testis-specific histone variant. Methylation is one of the main histone modifications: it is regulated by the enzyme methyltransferases and is generally associated with gene silencing [7]. Acetylation is regulated by acetyltransferase and deacetylates histones: the first one activates gene expression whereas the second inhibits it. Histone phosphorylation takes place on lysine residues and generally promotes gene activation [7]. Lastly, ubiquitination is a covalent modification that occurs on lysine residues and it is capable of activating or repressing gene expression [7].
Since retained histones are present in imprinted gene clusters [49], changes in protamination and in residual histones can be co-responsible for male infertility. Accordingly, a study carried out on 291 ART cycles described the role of histone-to-protamine ratio (HPR) on embryo development and ART outcome. For HPR ranging from 6 and 26%, the blastocyst formation rate was significantly higher (87.8%) compared to that obtained with HPR <6% (71.2%) or >6% (74.6%). Therefore, HPR seems to influence embryo development [59]. On the basis of such evidence, sperm devoid of protamine and residual histones abnormalities should be utilized in ART programs [49].
Go to :
SPERM RNA
Sperm carries thousands of different RNAs, including coding (mRNA) and non-coding [micro-RNA (miRNA), small interfering (siRNA), antisense RNA, piwiinteracting RNA (piRNA) and lncRNA] RNAs [60], the latter known to be involved in the modulation of gene expression, interfering with the mRNA translation through different mechanisms.
Overall, more than 4,000 different mRNAs have been detected in human spermatozoa [61]. Due to sperm chromatin compactness, it was thought that sperm RNA may unlikely be transcribed from sperm nuclear DNA and that it may be a residual of spermatogenesis [6263]. However, there is some evidence of transcriptional activity in spermatozoa [6465], thus suggesting that it may derive from imprinted genes mapping in unmethylated histones, where the transcriptional apparatus can access.
Sperm RNAs have been discovered in mammals, insects [66] and plants (pollen RNA [67]), suggesting that it might represent a conserved characteristic of the male gamete [68]. As far for sperm RNA functions, they are mostly completely unknown [666970]. Gathering together the available evidence from both animal and human studies, three main roles may be hypothesized: transmission of parentally acquired phenotype, spermatozoa maturation in the epididymis and embryo development, as discussed below.
Mounting evidence has shown that sperm RNAs are involved in the transmission of paternally acquired phenotypes [8]. Recently, the comprehensive profiling of sperm epigenome from obese and lean men resulted in a different sperm non-coding RNA expression, suggesting that sperm RNAs may act as a sensitive marker of environmental exposure [71]. Sperm RNAs are capable to modulate early embryonic gene expression [8]. Therefore, due to the higher risk of developing obesity which children of obese fathers are exposed to, the differential sperm RNA content of obese patients has been hypothesized as the possible mechanism through which obesity is inherited by the offspring [71].
Mammalian sperm RNAs are involved in sperm maturation [72,73]. This role might be displayed by piRNAs, that are 30 nucleotide-long non-coding RNAs interacting with the so called Piwi proteins. The Piwi/piRNA machinery acts in gene silencing that occurs in the male gamete [74]. During spermiogenesis, Piwi proteins seem to mediate the histone-to-protamine exchange in a piRNA-independent manner in mice. Piwi germline mutations preventing Piwi ubiquitination and degradation have been reported in azoospermic patients [75]. Despite further research is needed, the Piwi/piRNA machinery may be involved in human spermiogenesis, during the phase of late spermatids [75], probably acting on sperm DNA protamination.
Finally, sperm RNAs seem to be play a role in embryo development. Sperm RNAs are carried into the oocyte and, hence, translated [68]. A recent study performed in mice reported a significant decrease in blastocyst formation rate after ICSI when an RNase-treated sperm was used [76]. The treatment resulted in a 90% reduction of the sperm RNA, thus supporting that the latter is required for mice embryo development [76]. No study has explored the relationship between sperm specific transcripts and ART outcomes so far.
In conclusion, sperm RNAs may be involved in several important functions. The possible role that they may play in human fertilization and embryo growth needs to be elucidated.
Go to :
CONCLUSIONS
The study of the epigenetic pattern of spermatozoa can help to better understand causes of male infertility. Indeed, specific epigenetic aberrations can affect primary phases of embryonic development and ART outcomes. In particular, P1/P2 ratio, HPR and H19 DMR methylation rate may represent additional targets to investigate sperm quality before ARTs. Furthermore, sperm IGF2 and IGF1R expression should be further investigated.
In conclusion, imprinted unmethylated paternally-expressed genes in residual histones may be accessed from the transcriptional apparatus. It may be hypothesized that once transcribed, these genes contribute to the pool of sperm RNA. Abnormal DMR methylation rate of imprinted genes, alterations in chromatin compactness or in HPR represent possible mechanisms through which sperm RNA content, fertilization, embryo growth and, therefore, ART outcomes may be affected. Research characterizing sperm RNAs and their role in human fertility is needed.
Go to :
 XML Download
XML Download