

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Central nervous system (CNS) injury is an overwhelming event that can lead to neurological disorder with many sequelae. Due to the immune-privileged and complex nature of the CNS it was believed that the resolution of CNS injury is much more complicated and different from other mammalian tissue repair process. Most of the CNS injury victims develop irreversible disabilities like cognitive impairment, and even paralysis. In the sequelae of CNS injury, both in traumatic brain and spinal cord injury (SCI), the secondary injury following the primary insult is mostly responsible for the irreversible tissue damage [12]. Following CNS trauma, the impaired blood-brain barrier (BBB) alongside the brain-cerebrospinal fluid barrier, allows the cellular and chemical infiltration, which disrupts the microenvironment at the lesion site causing the acceleration of the cellular damage, degeneration, and scar formation [34]. The glial scar formation is believed to be the key player in forming the chemical and physical barrier for the neuronal regeneration, which impair the rehabilitation process after CNS injury [5].
Following CNS injury, glial cells play an important role in the injury and repair process. The glial cells astrocytes, microglia, oligodendrocytes, and their precursors interact with neuronal cells to maintain the cellular and extracellular microenvironments to exert basic CNS functions. Astrocytes are known to be the key provider of nutrients to the neurons and biochemical support such as ion homeostasis [6], neurotransmitter regulation [7], BBB maintenance [8], and the production of extracellular matrix molecules. They are also the most abundant cells in the mammalian CNS. Following CNS injury, astrocytes can sense the changes in the microenvironment and neuronal activity which influence the structural and functional changes in the astrocytes which eventually protects the brain or deteriorate the repair process [910]. Microglia, the brain-resident macrophages, surveilling the CNS environment in resting conditions, are central to the modulation of the inflammatory response. When they are activated following infection or injury, they can be transformed into activated form, and can secrete neurotoxic substances and molecules that magnify the immune/inflammatory responses, and retain phagocytic activity [111213]. Their function in CNS injury has some beneficial effects with respect to neuronal repair and inflammatory resolution. However, in many studies that blocked the innate immune response or anti-inflammatory treatment following CNS injury have found to restore the functional impairments [141516]. Oligodendrocytes and their progenitors provide myelin sheaths for wrapping the axons, which ensures proper axonal transmission and may also remyelinate in certain pathological conditions [171819]. Following CNS injury, the loss of the oligodendrocytes results in the loss of axonal myelination followed by impaired motor function and eventually paralysis.
Agmatine, a derivative of L-arginine produced via decarboxylation by arginine decarboxylase (ADC), has been mainly implicated in the study of modulation of neuronal functions [202122]. Agmatine interacts with different neurotransmitter receptors including the N-methyl-D-aspartate receptor, 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl) propanoic acid receptor, kainate receptor, acetylcholine receptor, serotonin receptor, α2-adrenoreceptor, and acid-sensing ion channels [23242526272829]. It has also been suggested that both exogenous and endogenous agmatine is involved in neuroprotection against various CNS injuries, including stroke and SCI. In this review, we will discuss the role of agmatine in the functional modulation of glial cells following CNS injury.
AGMATINE, NEUROGLIA, AND CNS INJURY
In any CNS injury glial cells play a vital role in the progression of the injury process and repair (Fig. 1). As a result, it is very important to modulate glial function following the onset of the injury until repair to ensure neuro-protection and accelerate the rehabilitation process. Both endogenous (mostly synthesised and stored in astrocytes [30]) and exogenous agmatine were found to be neuroprotective and modulate the function of glial cells.
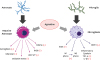 | Fig. 1Agmatine attenuates astrogliosis and microgliosis following CNS injury. Agmatine treatment reduces the detrimental effects of both reactive astrocytes and activated microglia after CNS injury. Agmatine treatment suppresses the M1 microglia and increases the M2 microglia via regulation of the BMPs expression. (−): suppression, (+): increase of expression.CNS, central nervous system; BMP, bone morphogenic protein; TNF, tumor necrosis factor; IL, interleukin.
|
Regulation of astrocyte function by agmatine in CNS injury
The most abundant cells in the mammalian CNS can be divided into two subclasses, astrocytes of the grey matter are known as protoplasmic and those in white matter known as fibrous astrocytes [31]. Originating from the neuroepithelial progenitor cells, astrocytes help in neuronal migration, growth, and development of neurons and synapses during mammalian development [32]. The only two diseases which have been found to be directly or partially related to astrocytes are astrocyte gene glial fibrillary associated protein defect and astrocyte associated hepatic encephalopathy [3334]. However, astrocytes play a very important role in the development and repair process of both chronic and acute diseases such as Alzheimer's disease (AD), Parkinson's disease, amyotrophic lateral sclerosis, Huntington's disease, epilepsy, stroke, SCI and others [353637383940]. In the CNS injury, astrocytes respond according to the location and severity of the injury. The post-injury expression of the pro-proliferative cytokines such as interleukin-6, transforming growth factor-α, ciliary neurotrophic factor, fibroblastic growth factor-2, and epidermal growth factor are known to increase astrocyte proliferation, leading to reactive astrogliosis and eventually formation of glial scar [41]. The reactive astrocytes present in the glial scar are well known to inhibit the regeneration of severed neurons and shunting neurite outgrowth by the inhibitory extracellular matrix, chondroitin sulphate proteoglycans (CSPGs) [4142]. The glial scar contributed CSPGs are also reported to be associated with the impairment of neural stem cell migration [43]. Intraperitoneal treatment of agmatine (100 mg/kg) (exogenous) has been found to reduce the severity of middle cerebral artery occlusion (MCAO) induced cerebral astrogliosis, which is also associated with increased neuroprotection via attenuating oxidative stress in rat [44]. To better understand the neuroprotective function of agmatine, Lee et al. treated agmatine in the in vitro primary astrocytes subjected to oxygen-glucose deprivation (OGD). Following OGD astrocyte viability was decreased, which was increased by the agmatine treatment through the nuclear translocation of NF-κB [45]. Agmatine also reduced the MCAO-induced brain oedema and overexpression of AQP4, a selective water-transporting protein highly expressed in the astrocytic foot process, which play an important role in brain water intoxication [46]. On the other hand, in the chlorpromazine (CPZ) induced rat forebrain injury, agmatine (75 mg/kg intraperitoneally [i.p.]) was found to accelerate astrogliosis which overlaps the increased expression of superoxide dismutase and reduction of the reactive oxygen [47]. In this case, the agmatine induced reactive astrocytes might exert their beneficial effect via suppression of the oxidative stress through lipid peroxidation. Agmatine was also found to stabilize the BBB, a principal function of astrocyte in rat transient cerebral ischemia [48]. However, the effect of agmatine on astrocytes on the contribution of BBB stabilization was not been reported in this study. Overexpression of endogenous agmatine via transduction of the human ADC gene was found to reduce oxidative stress injury in in vitro mouse cortical astrocytes [49]. In this study the authors suggested that that the neuroprotective function of endogenous agmatine were exerted via inhibiting inducible nitric oxide synthase (iNOS) signaling and metalloproteinases activity in astrocytes. In the mice SCI, it was also reported that agmatine treatment attenuated reactive astrogliosis and glial scar formation which was correlated with an increase in the level of bone morphogenic proteins 2/4/7 (BMP-2/4/7) [50]. BMP-7 has been suggested to be neuroprotective and a regulator of glial cell differentiation [5152]. Studies have also suggested that agmatine treatment can also suppress astrogenesis by suppressing BMP-2/7 expression in neuronal stem cells [53]. Thus, agmatine plays a vital role in the formation of reactive astrocytes and glial scar, which eventually modulate the repair and rehabilitation process after CNS injury. Further studies of the mechanism of action of agmatine on astrocytes are needed to ensure its proper therapeutic use in CNS injury.
Regulation of microgliosis by agmatine in CNS injury
The functional properties of the microglia in the healthy brain are still poorly understood. However, they are well known for their primary surveillance activity in the immune privileged brain, and are also considered to be the key player during the development, homeostasis, and modulation of CNS diseases. Microglia have the ability to scan the whole brain parenchyma within a few hours [54]. In the diseased or injured brain, microglia can be both beneficiary or detrimental to the brain by modulating disease progression and repair process. In AD, frontotemporal dementia, normal aging, axonal pruning has been reported to be associated with an increase in microglial expression of C1q, accompanied by compliment component C3 [555657]. In damaged tissue, microglia promote the induction of astrocytes associated proinflammatory cytokine expression, such as IL-1α, tumor necrosis factor, and C1q which lead to expression of C3. C3 is considered to be cytotoxic to neurons and oligodendrocytes [58]. CNS injury also induces an astrocyte associated ATP gradient, which is sensed by the microglia through purinergic receptor P2RY12, leading to rapid changes in the microglia and increased microglial migration to the injury site [5960]. Recent studies have suggested that microglial cells can be present as one of the two phenotypes according to their activation and functional modalities such as either classically activated (M1) or alternatively activated (M2) [6162]. Among them, M2 microglia are suggested to be beneficial following the CNS injury due to the suppression of inflammation, promoting tissue repair, and wound healing. Following CNS injury, M2 microglia are believed to be express at earlier stages of injury which become pick at 5th day of injury and are later converted to M1 phenotype [6364]. M1 microglia are suggested to increase the iNOS expression, which promotes neurotoxic NO production in in vitro ischemic condition [63]. In the BV2 microglial cell-mediated in vitro model of the ischemic condition, agmatine (100 μM) treatment was found to attenuate the cytotoxic condition and NO production was correlated with the reduction of iNOS production [65]. The authors also suggested that agmatine (100 mg/kg i.p.) treatment reduced the expression of ionized calcium binding adaptor molecule 1 (Iba1), which is a well-known marker for microglia and iNOS in the in vivo MCAO rat model. The same research group also found the same effect of agmatine treatment in both in vitro and in vivo (mice) model of lipopolysaccharide (LPS) induced inflammatory injury [66], which supports the results of the LPS-induced iNOS expression and NO production in rat primary cortical microglia [67]. Moreover, they suggested that agmatine can attenuate the expression of pro-inflammatory cytokines, such as tumor necrosis factor-α and interleukin-1β, induced by the LPS-injured brain [66]. In the CPZ treated oxidative stress model of wistar rats' brain, agmatine reduced the expression of Iba1 [46]. In rat SCI model, agmatine was found to be a modulator of BMP-2 expression along with the reduced expression of M1 macrophages which was increased following injury, and increased M2 expression [20]. Recently, BMP-2 has been reported to be a suppressor of M1 macrophages [68]; based on the above results, the authors suggested that agmatine is responsible for the transition of M1 macrophage to M2 via increased BMP-2 expression. Although, agmatine has been found to modulate CNS injury and positively accelerate the repair process, more studies are needed to confirm the therapeutic role of agmatine on microglia and macrophage.
Agmatine promotes myelination by oligodendrocytes in CNS injury
Oligodendrocytes are the end product of the cell linage with a complex timed program including proliferation, differentiation, migration and later myelination. They are also known as the major myelinating cells, which produces cholesterol myelin sheaths to insulate axons to ensure and accelerate the proper axonal transduction process [6970]. The role of the myelin is not only to ensure axonal signal transduction, but in CNS diseases it may underlie severe functional and rehabilitation disabilities [7172]. Loss of myelination may happen in different types of diseases or injuries such as genetic, infectious and autoimmune disease, and also following CNS trauma [737475]. There are not many studies has been found that examine the role agmatine on oligodendrocytes. However, Park YM. et al. suggested that exogenous agmatine treatment promoted remyelination following compression induced SCI in a mouse model that was correlated with the functional neurobehavioral outcome (Fig. 2) [49]. They also suggested that agmatine treatment increased the expression of the Olig-2+ oligodendrocyte cells and also prevented the loss of neuronal cells. The expansion of oligodendrocytes from NG+ oligodendrocyte progenitor cells has been reported to be regulated by the higher expression of BMP-2/7 and decreased BMP-4 expression influenced by agmatine [49]. The transplantation of human mesenchymal stem cells overexpressed with the endogenous agmatine-producing gene, ADC, in SCI reduced glial scar formation and promoted neuronal regeneration and oligodendrogenesis [76]. Braissant et al. [77] suggested that the blood to cell exchanger of L-arginine, a substrate of agmatine, cationic amino acid transporters (CAT2) is expressed in neuron and oligodendrocyte, which suggests that the agmatine might be closely related to oligodendrocytes. These results warrant future studies examining the role of agmatine on oligodendrocytes.
 | Fig. 2AGM increases remyelination following CNS injury. (A) AGM treatment increases the remyelination (luxol blue staining) 28 days after SCI in the rat, which is also correlated with the significantly increased functional outcome compared to experimental control (B). Results represent mean ± standard error of the mean.CNS, central nervous system; BMS, basso mouse scale; EC, experimental control; NC, normal control; AGM, agmatine.
*p < 0.05, EC vs. AGM treated group.
|
CONCLUSION
Agmatine has been found to have neuroprotective effect in both acute and chronic CNS disorders. Agmatine modulates different receptors and ion channels that are responsible for the progression and resolution of CNS injury. There are a number of studies that reported that, both endogenous and exogenous agmatine possess beneficiary role on most CNS cells such as neuron, astrocytes, microglia and oligodendrocytes and also modulate the differentiation, proliferation and migration of those cells. Therefore, it is important to understand the mechanism of agmatine action on CNS cells which will help to understand the therapeutic potential of agmatine in CNS injury.
 XML Download
XML Download