

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
An entire view of human intracardiac nerves seemed to be first provided by Pauza et al. [1] who used an excellent technique of whole-mount histochemistry of acetylcholinesterase for human fetal hearts, they demonstrated that (1) epicardiac nerves provide seven subplexuses and (2) more than half ganglion cells are present at the dorsal and dorsolateral surfaces of the left atrium. They also ensured a specifically concentrated nerve bundles in a fold separating the entrance of the left superior pulmonary vein from the left atria, i.e., the so-called left atrial nerve fold. This fold was first reported by Worobiew [2] and Gardner and O'Rahilly [3] later ensured the presence. This fold seems to correspond to the future atrial-pulmonary vein junction, the latter of which is likely to contain a histopathological substrate of chronic atrial fibrillation [4567]. In addition to whole-mount studies, several histological and immunohistochemical studies had been conducted using sections of human adult hearts, not of fetuses [89101112]. In these reports, Marron et al. [8] was characterized by detailed descriptions of nerve terminal morphologies but, to our regret, there was no information about the atrial and ventricular septum.
According to our experience [13], the whole-mount histochemistry has a disadvantage that, in contrast to surface nerves, nerves embedded deeply in a connective tissue are not clearly demonstrated or even not stained because of limited infiltration of substrates for reaction. Likewise, a nerve running through a matured muscle is also difficult to demonstrate because of difficult transillumination through the dark colored tissue. Thus, even in beautiful photographic by Pauza et al. [1], nerves in and through the ventricular myocardium were not demonstrated. Moreover, histological studies using human fetuses had focused on extrinsic innervation to the sinoatrial and atrioventricular nodes [314]. Therefore, especially in the late stage fetuses with thick and matured cardiac muscles, there was no or few morphological information on (1) nerves around the atrioventricular orifices and and (2) nerves running through the ventricular wall and septum. The late stage fetus has also an advantage that the atrial-pulmonary vein junction is not yet established in association with a widely opened oval foramen. S100 protein was a neurospecific protein. This protein was generally considered to be a component in the glial and Schwann cells of peripheral nervous system [15]. Consequently, using histological sections with S100 protein immunohistochemistry, the aim of this study was to examine intracardiac nerve configuration in the late stage fetuses.
Go to : 
Materials and Methods
This study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in 2013). We used frontal or sagittal sections of 12 masses of thoracic viscera obtained from 12 human fetuses at 25–33 weeks (crown-rump length, 200–280 mm). These fetuses were parts of a collection in Department of Anatomy, Akita University, Akita, Japan. They were donated by their families to the department during 1975–1985 and preserved in 10% w/w neutral formalin solution for more than 30 years. The available data was limited to the date of donation and the gestational weeks, but we did not find a document saying the family name, the name of obstetricians or hospital and the reason of abortion. The use for research was approved by the university ethics committee in Akita (No. 1428).
We prepared 12 paraffin blocks containing the whole heart as well as parts of the lung, thymus and diaphragm. From each of the blocks, we prepared 200-300 semiserial sections covering the entire heart (5-µm in thickness; 50-µm interval). One of every 5–10 sections were used for immunohistochemistry of S100 protein, while the others were used for stained with hematoxylin and eosin. The primary antibodies used were and a rabbit polyclonal anti-human S100 protein or S100 (dilution 1:100, Dako N1573, Dako, Glostrup, Denmark). After incubation for 30 minutes in Histofine Simple Stain Max-PO (Nichirei, Tokyo, Japan) for the diaminobenzidine (DAB) reaction with horseradish peroxidase, dark brown coloration (DAB reaction) were obtained. Sections stained using the DAB method were counterstained with hematoxylin. Observations and taking photographs were usually performed with Nikon Eclipse 80, but photos at ultra-low magnification (objective lens less than ×2) were taken using a high-grade flat scanner with translucent illumination (Epson scanner GTX970, Java, indonesia).
Go to :
Results
Intracardiac nerves and ganglia are demonstrated (1) at and along the atrial wall (Figs. 1, 2, 3, 4) including the left atrial nerve fold (Fig. 2C, D) and the atrial septa (Figs. 3E, 4C–F), (2) in the ventricular wall including the ventricular septa and annuluses of the mitral and tricuspid valves (Figs. 2B–E, 3B, 4B, 5) and (3) in the atrioventricular sulcus and at the base of great arteries (Figs. 1A, B, 6). Each of the figures is prepared for a single fetus except for Figs. 3 and 4 those display the primary and secondary atrial septa (septum primum and septum secundum) of the same specimen.
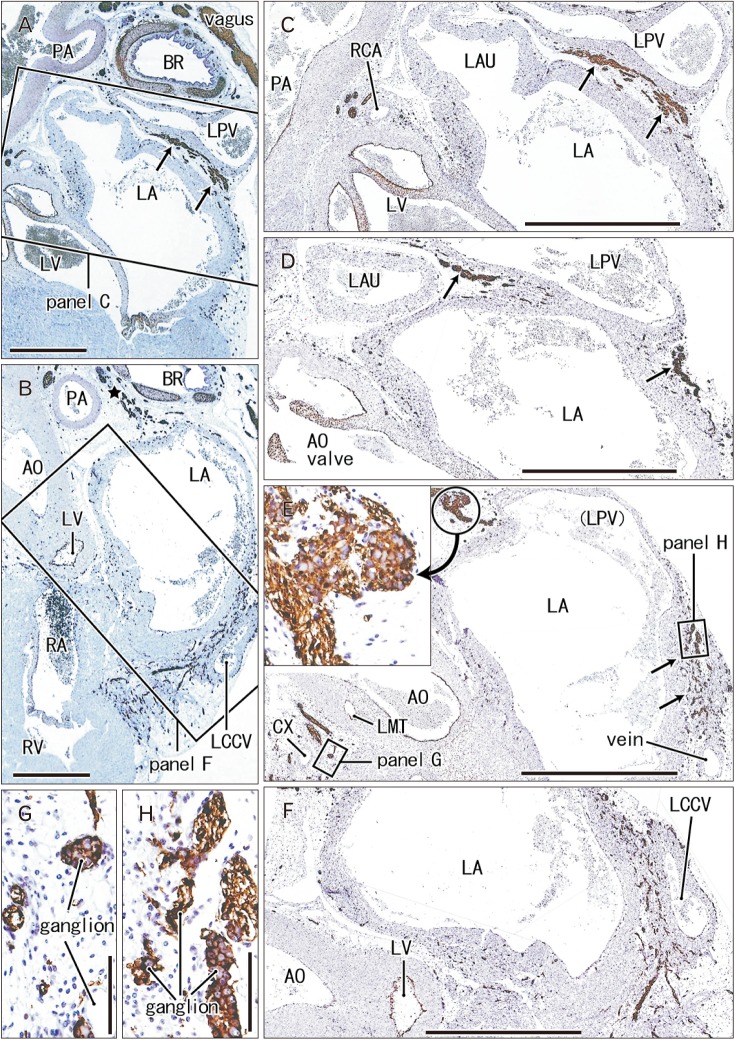 | Fig. 1Nerves descending between the left atrium (LA) and pulmonary vein to a site along the left common cardinal vein. 25 weeks. Sagittal sections. Immunostaining of S100 protein. Incomplete square in panel A (or B) indicates an area of panel C (or F) at the higher magnification. Ganglia in panel E are shown in panels G and H as well as in an insert at the center of this figure. Panels D and E display intermediate planes between panels C and F. The left pulmonary vein (LPV) drains into the LA in a plane between panels D and E. Abundant nerves around the left common cardinal vein (LCCV in B and F) comes from a superior site (star in B) between the left primary bronchus (BR) and pulmonary artery (PA) via a narrow space between the LPV and LA (arrows in panels A, C–E). AO, aorta; CX, circumflex branch of the left coronary artery; LAU, left auricle; LMT, main trunk of the left coronary artery; LV, left ventricle; RA, right atrium; RCA, right coronary artery; RV, right ventricle; Vagus, left vagus nerve. Scale bars=5 mm (A, B), 1 mm (C–F), 0.1 mm (G, H).
|
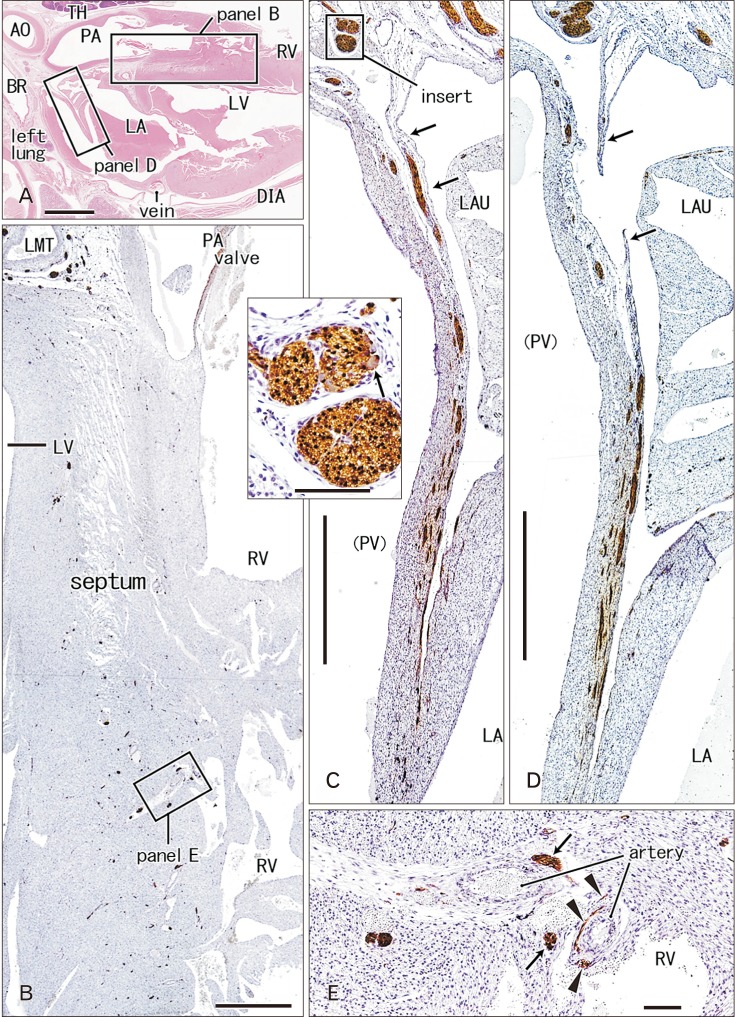 | Fig. 2Nerves and ganglia in the left atrial nerve fold and interventricular septum of the heart. 26 weeks. Sagittal sections through the outflow tract of the right ventricle (RV) and the interventricular septum. Hematoxylin and eosin staining (A) and immunostaining of S100 protein (B–E). Squares in panel A are shown in panels B and D at the higher magnification. Panel B shows abundant thin nerves in the interventricular septum: parts of muscular nerves (arrows in E) as well as nerve nets around arteries (arrowheads in E) are exposed to the expanding ventricular endothelium. Panel D as well as panel C (a plane 1 mm left side of D) exhibits the left atrial nerve fold: most of the nerve contents run through the future wall of the pulmonary vein (PV) and they come through a pericardial fold (arrows). An insert between panels B and C displays a ganglion cell (arrow) at the base of the nerve fold. AO, aorta; BR, bronchus; DIA, diaphragm; LA, left atrium; LAU, left auricle; LMT, left main trunk of the coronary artery; LV, left ventricle; PA, pulmonary artery; TH, thymus. Scale bars=5 mm (A), 1 mm (B–D), 0.1 mm (E, insert between B and C).
|
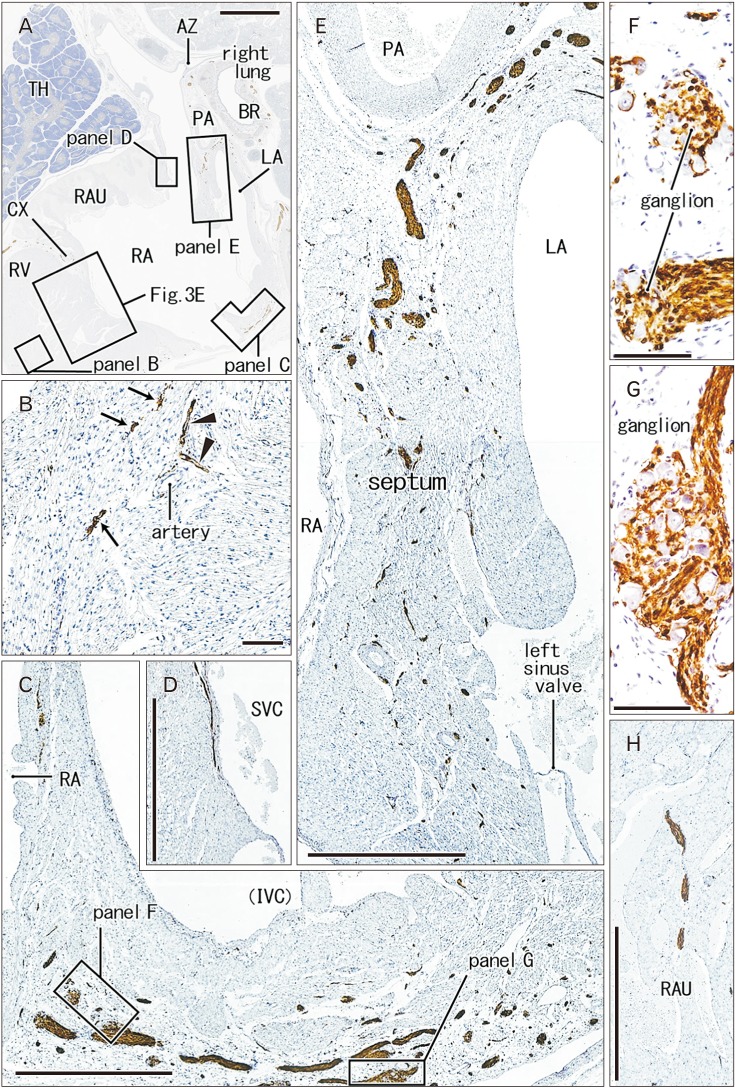 | Fig. 3Nerves and ganglia in the right ventricle and atrium of the heart. 31 weeks. Sagittal sections. Immunostaining of S100 protein. Squares in panel A are shown in panels B–E as well as Fig. 3E at the higher magnification. Panel B displays muscular nerves (arrows) and nerve nets along an artery (arrowheads) in the right ventricular wall. Panel C exhibits the right atrial wall near the orifice of the inferior vena cava (IVC): the ganglia are shown in panels F and G at the higher magnification. Panel D displays a nerve along the endothelium of the superior vena cava (SVC). Panel E shows nerves in the interatrial septum near the orifice of the pulmonary vein. Panel H, a plane 3 mm right of the other panels, exhibits a thick nerve passing through the right auricle (RAU). AZ, azygos vein; BR, bronchus; LA, left atrium; PA, pulmonary artery; RA, right atrium; RV, right ventricle; TH, thymus. Scale bars=5 mm (A), 0.1 mm (B, F, G), 1 mm (C–E, H).
|
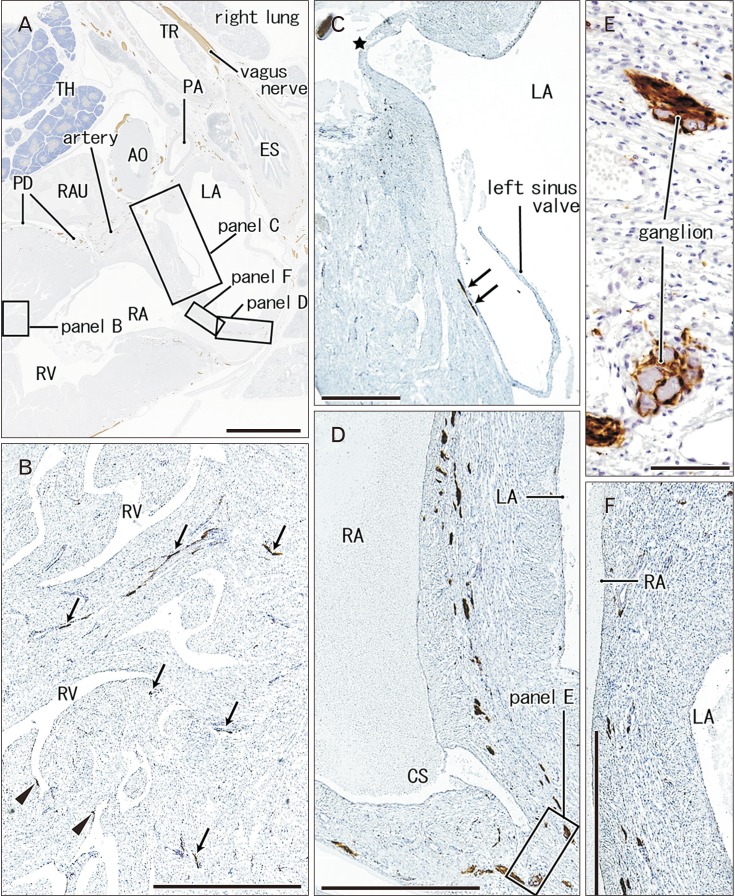 | Fig. 4Nerves and ganglia in the interatrial septum and right ventricular wall. 31 weeks. A specimen same as in Fig. 5. Sagittal sections 3 mm medial or left side of Fig. 4. Immunostaining of S100 protein. Squares in panel A are shown in panels B–D and F at the higher magnification. Panel B displays nerves (arrows and arrowheads) in the right ventricular wall: some of them (arrowheads) are exposed to the ventricular endothelium. Panel C exhibits the upper part of the interatrial wall, while panels D and F the lower part. Panel C contains a nerve along the endocardium (arrows) as well as an orifice of a minor cardiac vein draining into left atrium (star). Ganglia near the cavernous sinus (CS) are shown in panel E at the higher magnification. AO, aorta; ES, esophagus; LA, left atrium; PA, pulmonary artery; PD, posterior descending branch of the right coronary artery; RA, right atrium; RAU, right auricle; RV, right ventricle; TH, thymus; TR, trachea. Scale bars=5 mm (A), 1 mm (B–D, F), 0.1 mm (E).
|
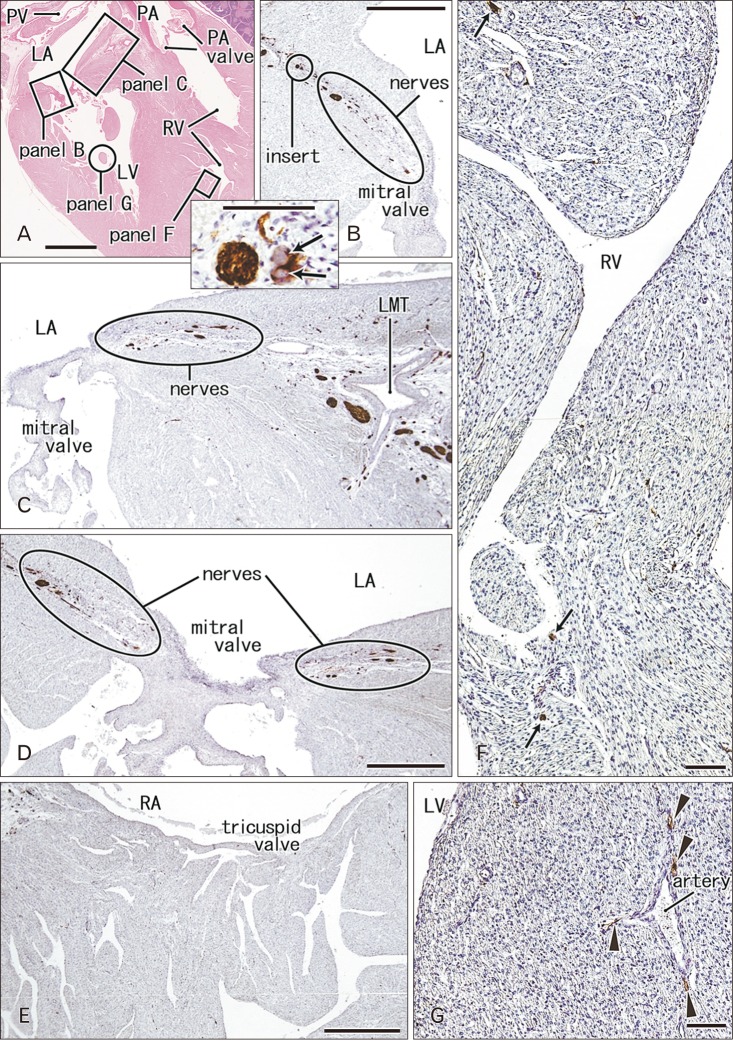 | Fig. 5Nerves and ganglia around the atrioventricular valves and in ventricular muscles of the heart. 31 weeks. Sagittal sections through the outflow tract of the right ventricle (RV) and the mitral valve of the left ventricle (LV). Hematoxylin and eosin staining (A) and immunostaining of S100 protein (B–G). Squares in panel A are shown in panels B, C, F and G at the higher magnification. Panels B–D display nerves around the mitral valve annulus. Panel D shows a plane 0.5 mm left side of panel C. An insert between panels A and B displays ganglion cells (arrows) near the base of the mitral valve. Panel E exhibits the tricuspid valve of the same specimen (corresponding to a square in Fig. 4A). In contrast to abundant nerves around the mitral valve annulus (B–D), few nerves are seen around the tricuspid valve annulus (E). Panel F shows muscular nerves (arrows) in trabeculae of the RV: parts of them are exposed to the expanding ventricular endothelium. Panel G displays a nerve net (arrowheads) along an artery in the papillary muscle of the LV. LA, left atrium; LMT, left main trunk of the coronary artery; PA, pulmonary artery; PV, pulmonary vein; RA, right atrium. Scale bars=5 mm (A), 1 mm (B–E), 0.1 mm (F, G, insert between A and B).
|
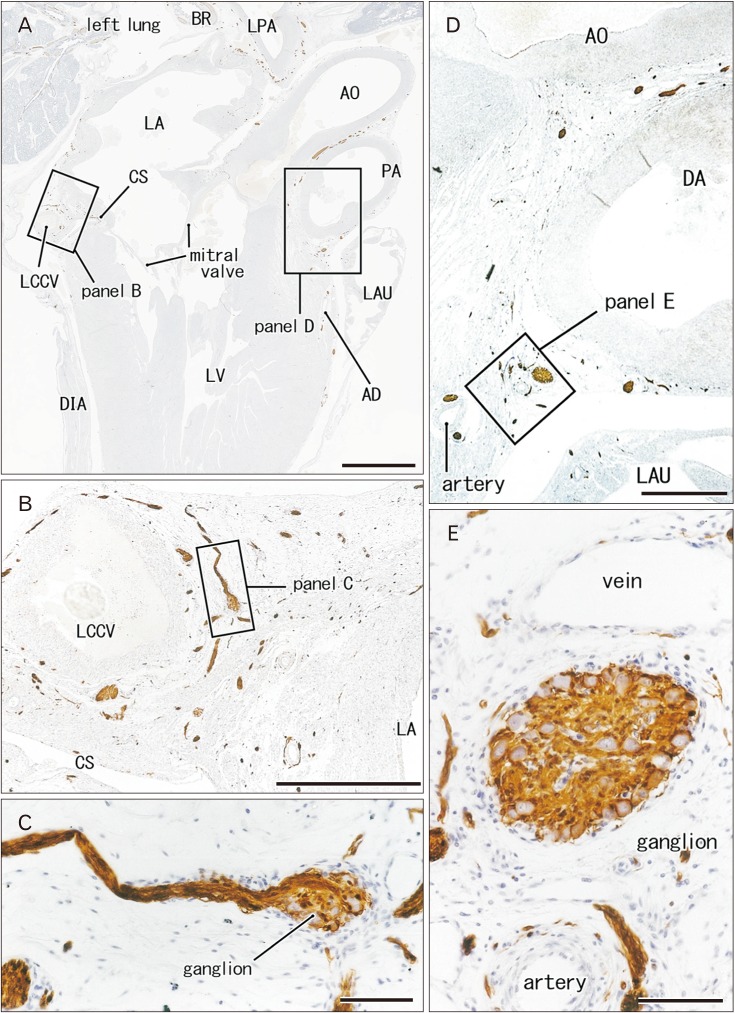 | Fig. 6Nerves and ganglia along the coronary sulcus of the heart. 30 weeks. Sagittal section through the aortic outflow tract of the left ventricle (LV) as well as the mitral valve. Immunostaining of S100 protein. Two squares in panel A are shown in panels B and D at the higher magnification. A ganglion in panel B or D is shown in panel C or D at the higher magnification. The left common cardinal vein (LCCV) is degenerating and obliterated in a plane 3 mm left of panel B, but it accompanies abundant nerves (B). AD, anterior descending branch of the left coronary artery; AO, aorta; BR, bronchus; CS, coronary sinus; DA, ductus arteriosus; DIA, diaphragm; LA, left atrium; LAU, left auricle; LPA, left pulmonary artery; PA, pulmonary artery. Scale bars=5 mm (A), 1 mm (B, D), 0.1 mm (C, E).
|
Ganglion cell bodies were round or oval and identified as negative areas in S100 immunohistochemistry. They ranged from 10 to 30 µm in diameter and the size was smaller in the earlier or smaller specimens, e.g., Fig. 1 (25 weeks) vs. Fig. 6 (30 weeks). Ganglion cell clusters were distributed (1) along the atrioventricular sulcus especially near the crux (Figs. 1E, 3C, 4D, 6B), (2) along the left main trunk of the left coronary artery (Figs. 1E, 5C, 6B), (3) at the origin of the right coronary artery (Fig. 1C), and (4) in front of the pulmonary arterial trunk (Fig. 6D). These ganglionated nerve plexuses corresponded to the left and right coronary subplexuses of Pauza et al. [1]. When roughly estimated using present semiserial sections with 50-µm interval, numbers of ganglion cell bodies ranged from 170 to 250 between the ascending aorta and pulmonary trunk, 150–200 at and near the crux and 30–150 in front of the pulmonary trunk.
The left common cardinal vein had lost the vascular lumen near the posterior body wall due to degeneration. This vein accompanied abundant nerves in and outside of the atrioventricular sulcus (Figs. 1F, 6B). These nerves came from an extracardiac nerve plexus between the left primary bronchus and pulmonary artery (Fig. 1A) via a narrow space between the left pulmonary vein and left atrium (Fig. 1C, D). They were likely to correspond to the left and middle dorsal subplexuses of Pauza et al [1]. Likewise, abundant nerves were also evident along the left superior pulmonary vein as well as a site between the vein and left atria (Fig. 2C, D): the vein and atria made a thick and wide fold corresponding to the left atrial nerve fold (see the first paragraph of the Introduction). These nerves appeared to correspond to the ventral left atrial subplexus and/or the left dorsal subplexus of Pauza et al. [1]. Most nerves to the left atrial nerve fold came from an extracardiac nerve plexus under the aortic arch as well as between the arch and ductus arteriosus (Fig. 2A). This nerve origin usually contained a large ganglion (ganglion cardiacum) near the origin of the left recurrent laryngeal nerve.
In the ventricular myocardium including the atrioventricular junction and interventricular septum, nerves were distributed deeply and seen with or without vessels accompanied (Figs. 2B, 3B, 5G). Notably, a high density of nerves was consistently evident around the mitral valve annulus, whereas the tricuspid valve annulus accompanied much less numbers of nerves (Fig. 5C–E). As ventricular myocardium was degenerated in the luminal side for sculpturing of papillary muscles and trabeculae, nerves near the ventricular cavity became exposed to the ventricular cavity (Figs. 2E, 4B, 5F). Likewise, as pectineal muscles were sculptured, nerves were exposed in the atrial cavity (Fig. 3H). Sometimes (3 specimens), we found thin nerves running along a long distance (0.5–1.0 mm) beneath the atrial endothelium (Figs. 3D, 4C).
In the posterosuperior and left sides of the oval foramen, the primary septum was attached tightly to the secondary septum. The attaching or fusion of the septa varied between specimens in area. The upper tip of the primary septum appeared to be a thin fold or valve arising from the original septum (Fig. 4C). The valve of the oval foramen carried no nerves and vessels. Notably, the primary septum contained few nerves and no vessels, whereas the secondary septum had abundant nerves and vessels (Figs. 3E, 4D, F). The myocardium along and near the coronary sinus and inferior caval orifice contained abundant nerves and ganglion cells (Figs. 3C, F, G, 4D, E).
Go to :
Discussion
The most striking feature in the present study was found in the luminal aspects of the ventricular and atrial wall: early-developed nerves became exposed to the ventricular or atrial endothelium due to sculptured myocardium during development of papillary muscles, pectinate muscles and trabeculae. Remodeling of myocardium occurs after midterm [16], partly involves cell death [17] and regulated by bone morphogenetic protein 4 (BMP4) and fibroblast growth factor 2 (FGF2) [18]. The latter group demonstrated that, in vitro, BMP4 induces, whereas FGF2 inhibits, apoptosis of mouse embryonic ventricular myocardium. The process to provide papillary and pectinate muscles was previously termed “luminal trabeculation” and recently called “myocardial assembly” [19]. Possibly due to unbalance between luminal expansion and myocardial growth, parachute-like asymmetric growth occurs for development of papillary muscles [20]. Therefore, conversely, early nerve growth in the myocardium seemed to be out of the regulation for myocardial assembly.
We found another striking feature in the growing atrial septum: the secondary septum contained abundant thick nerves in contrast to no or few nerves in the primary septum. This difference was most evident at the left posterior margin of the oval foramen in which these two septa are attached tightly. Singh et al. [21] found several ganglion cells in the adult atrial septum, but we did not in fetuses. Thus, the postnatal development of ganglion cells seemed to be likely in the atrial septum. We also found a clear difference in nerve density between the mitral and tricuspid annuluses: a high density of nerves around the mitral valve in contrast to few nerves around the tricuspid valve. This difference might be connected with arrhythmia after replacement surgery of these valves. However, a patient requiring tricuspid valve replacement is usually suffering from a combination of anomalies such as Ebstein's anomaly [2223]. Thus, it seemed to be difficult to compare between clinical outcomes of the mitral and tricuspid valve surgeries
According to our recent study [24], epicardiac nerves and ganglion cells along the left common cardinal vein are quite different from those at the origins of the great arteries in proportion of tyrosine hydroxylase (TH)- and neuronal nitric oxide synthase (nNOS)-positive neurons. The present observations suggested that, depending on expansion of the left atrium, the nerves along the left common cardinal vein were likely to be involved in the atrial-pulmonary vein junction. Likewise, nerves in the left atrial nerve fold were also likely to supply the atrial-pulmonary vein junction because the growing fold made the atrium and vein more separated. These nerves came from the ganglion cardiacum and this ganglion contains abundant TH-positive neurons and a few nNOS-positive neurons. Therefore, nerves to the atrial-pulmonary vein junction seemed to have dual origins and fiber components. Pauza et al. [1] considered that the left atrial fold nerve is originated from nerves passing through the venous pole of the heart: this was not contradict to the present observations since parts of the composite nerve fibers came along the left common cardinal vein. The atrial-pulmonary vein junction is likely to contain a histopathological substrate of chronic atrial fibrillation: the pathological difference in substrate is likely to be found in phenotypes of neurons [457], but the other researchers considered it simply as a difference in nerve density [625]. A proportion between the dual nerve origins in fetuses might be connected with an individual difference in histological substrate.
Go to :
 XML Download
XML Download