

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Scrapie, a transmissible neurodegenerative disease in sheep and goats, is the oldest known animal prion disease [1]. There are many strains of scrapie, distinguishable by their disease characteristics with each strain known to exhibit tropism targeting specific brain regions. The ME7 scrapie strain was originally isolated from a scrapie-infected sheep in the Suffolk sheep flock of the Moredun Research Institute in the UK [2]. The mouse-adapted ME7 scrapie strain serially passaged in C57BL/6J mice induces neurodegeneration and gliosis in the hippocampal brain region of infected animals. Proteinase K (PK) resistant scrapie prion protein (PrPSc) is readily detected in the late stage of scrapie and is widespread throughout the hippocampus [3]. Furthermore, ME7-infected animals have been shown to display marked vacuolation of the hippocampal, septal, and thalamic neuropils.
In the present study, we aimed to confirm the re-transmissibility of the mouse-adapted ME7 strain to ovine prion protein (PrP) transgenic mice and to describe the changes that might accompany the re-transmission. We report that interspecies transmission of the mouse-adapted ME7 scrapie strain to transgenic mice expressing ovine PrP was successful and it was accompanied by biophysical (glycoform pattern change) and neuropathological (PrPSc deposition with plaque formation) changes.
MATERIALS AND METHODS
Animals and scrapie infectious agent inoculation
The ovinized transgenic mouse line carrying the Suffolk sheep PrP gene (TgSShpPrP) containing a Kozak ribosome-binding sequence was generated [4] and kindly provided by Dr. Richard I. Carp (New York State Institute for Basic Research in Developmental Disabilities, USA). The C57BL/6J mice were purchased from Jackson Laboratory (USA). The prion strain ME7 was a gift from Alan Dickson (Neuropathogenesis Unit, UK) and was maintained by serial intracerebral passage of brain homogenates from terminally affected mice. ME7-infected brain tissues at the terminal stage of the disease were homogenized in 0.01 M phosphate-buffered saline (PBS) under aseptic conditions using a sterile disposable instrument (Precision tissue grinder system, USA). Six-week-old female TgSShpPrP mice were anesthetized by isoflurane inhalation and inoculated with 30 µL of 1% (w/v) brain homogenates through an intracerebral route. Control inoculum was prepared from the brain of uninfected TgSShpPrP mice. Post-inoculation, the mice were observed daily until neurologic symptoms developed. Animals were sacrificed when ataxia, altered gait, hind limb weakness, and leg-clasping reflex were noticed. Brains of both control and infected mice were harvested for protein assay and histological pathology of prion disease. All animal experiments were approved by the Hallym Medical Center Institutional Animal Care and Use Committee (HMC-2018-0-0521-7), Korea. The procedures were conducted in a manner that minimized animal suffering and the number of animals used.
Western blot analysis
ME7-infected TgSShpPrP mouse brains were homogenized with 10% volumes of radio immunoprecipitation assay buffer (Thermo Fisher Scientific, USA) containing complete protease inhibitor cocktails (Roche, Germany). The brain homogenates were centrifuged at 13,500 r/min for 15 min at 4°C. Aliquots of supernatant frozen at −70°C were used for western blotting. Protein concentration was determined by bicinchoninic acid assay (Thermo Fisher Scientific). To detect PrPSc, 7µg/mL of PK was treated for 1 h at 37°C. Equal amounts of protein (20 µg) were heated to 95°C for 5 min before being loaded into each lane in 15% sodium dodecyl sulfate (SDS) gel with a loading buffer containing 0.125M Tris-HCl (pH 6.8), 20% glycerol, 5% SDS, 10% mercaptoethanol, and 0.002% bromophenol blue. The separated proteins were transferred to nitrocellulose membrane (Amersham, USA) using an electrophoretic transfer system (BioRad, USA) at 90 V for 2 h. The membranes were washed with a Tris-buffered saline solution (pH 7.6) containing 0.05% Tween 20 (TBST) and then blocked in TBST containing 5% skim milk (Santa Cruz Biotechnology, USA) for 60 min at room temperature. The membranes were then incubated at 4°C overnight with one of the following antibodies: mouse monoclonal anti-PrP antibody (7A12, 1:5000), rabbit polyclonal anti-glial fibrillary acidic protein (GFAP) antibody (1:10000, Dako, Denmark), mouse monoclonal anti-NeuN antibody (1:1000, Millipore, USA). After washing in TBST, membranes were incubated with horseradish peroxidase-conjugated secondary antibodies against mouse and rabbit immunoglobulin G (IgG) (Sigma-Aldrich, USA) for 1 h at room temperature and washed in TBST again. The target signals were visualized using digital images captured with an Image Quant TM LAS 4000 imager (GE Healthcare Life Sciences, USA) and an enhanced chemiluminescence western blot detection system (Thermo Fisher Scientific). Western blot analysis (n = 3 for each group) was performed at least three times.
For peptide N-glycosidase F (PNGase F) treatment, brain preparations were treated with 0.5 µg/mL of PK for 1 h prior to PNGase F treatment. PNGase treatment was performed using a PNGase F enzyme kit (P0704S; New England BioLabs, USA) as recommended by the supplier. Reaction mixtures were precipitated with 1:1 methanol/acetone overnight at −20 °C and centrifuged at 14,000 r/min for 1 h. The obtained pellets were then subjected to immunoblotting.
Immunohistochemistry
To detect PrPSc, immunohistochemical analyses were performed using an ABC kit (Vector Laboratories, USA) according to a modified avidin-biotin-peroxidase method [5]. Briefly, 5-μm thick paraffin-blocked brain sections were deparaffinized with xylene, hydrated in a graded ethanol series, and then treated with 10 mM sodium citrate (pH 6.0, 0.5% Tween 20) for an antigen retrieval reaction in a microwave oven (LG Electronics, Korea) for 15 min. To stain the PrP, 10 µg/mL of PK were treated for 7 min at room temperature. After three washes in PBS, the tissue slides were treated with 0.3% hydrogen peroxidase in methyl alcohol for 30 min to block endogenous peroxidase activity at room temperature. Non-specific sites were blocked with blocking buffer (10% normal goat serum, 0.1% BSA, 0.1% Triton X-100 in PBS) for 1 h at room temperature. Tissue slides were then incubated overnight at 4 °C with anti-PrP antibody 3F10 (1:100) [5]. After rinsing with PBS, tissue slides were treated with biotinylated goat anti-mouse IgG (1:500) at room temperature for 1 h and avidin-biotin-peroxidase complexes (Vector Laboratories) and developed with a diaminobenzidine (DAB)-hydrogen peroxide solution (0.0005% DAB and 0.03% hydrogen peroxide in 50 mM Tris buffer). Next, the tissue sections were counterstained with Harris Hematoxylin solution, Gill No. 3 (Sigma, USA) and examined under a light microscope (Olympus BX51; Olympus, Japan). For immunofluorescence staining, brain sections were incubated with mouse monoclonal anti-NeuN antibody (1:200, Millipore) and rabbit polyclonal anti-GFAP antibody (1:200, Dako). Sections were subsequently visualized with Alexa Fluor 488-conjugated and 568-conjugated goat anti-mouse and anti-rabbit IgG (Molecular Probes, USA). Fluorescence images were obtained under a confocal laser staining microscope using Zen software (LSM-700; Carl Zeiss, Germany). In addition, double-labeling was performed to examine the distributions of neurons and astrocytes.
Semi-quantitative grading for lesion profile
Semi-quantitative vacuolar degeneration grading was performed in nine standard sites of each mouse brain by applying previously established methods [6]. Briefly, areas of vacuolar degeneration in mouse brain tissue sections were counted under a light microscope (BX51, 100 × magnifications; Olympus). Three tissue sections from each of the three tissue blocks for each mouse were used for lesion counting. The scoring system assessed the number and size of vacuolar changes and was expressed by the following scale: 0, no vacuolation; 1, a few vacuoles widely and unevenly scattered; 2, a few vacuoles evenly scattered; 3, a moderate number of vacuoles evenly scattered; and 4, marked vacuolation [7].
RESULTS
Serial transmission of mouse-adapted ME7 scrapie strains to TgSShpPrP mice
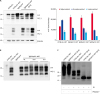
Intracerebral inoculation of the mouse-adapted ME7 scrapie strain to transgenic mice expressing ovine PrP resulted in prion diseases with distinct clinical manifestations. In this study, the incubation period for ME7-infected C57BL/6J mice was 148.0 ± 1.19 days while that of ME7-infected TgSShpPrP mice at first passage was 227.0 ± 6.35 days, with reductions to 185.3 ± 5.41 days and 132 ± 1.55 days at 2nd and 3rd passages, respectively (Fig. 1).
Fig. 1
Survival curve of TgSShpPrP mice. Comparative survival days of TgSShpPrP mice inoculated with a mouse-adapted ME7 scrapie strain at 1st, 2nd, and 3rd passages. The abscissa shows the survival days and the y-axis shows the percentages of survival. The average survival days and number of animals used are as follows: ME7-C57BL/6J, 148 ± 1.19 (n = 9); ME7-TgSShpPrP (1st), 227 ± 6.35 (n = 13); ME7-TgSShpPrP (2nd), 185 ± 5.41(n = 11); ME7-TgSShpPrP (3rd), 132 ± 1.55 (n = 5).
PrP, prion protein; TgSShpPrP, transgenic mouse line carrying the Suffolk sheep PrP gene.

Differential glycoform patterns of the abnormal PrP in the brains of C57BL/6J and TgSShpPrP mice infected with ME7 scrapie strain
We analyzed brain tissues from ME7-infected C57BL/6J and TgSShpPrP mice to determine if distinct abnormal PrP profiles capable of differentiating the disease phenotypes in the two species of mice could be identified. Immunoblotting was done using PK enzyme-treated brain homogenates obtained from control, C57BL/6J-ME7, and TgSShpPrP-ME7 mice. We expected that the PrPSc glycoform pattern in C57BL/6J-ME7 mice would be the same as that for TgSShpPrP-ME7 mice, especially when the ratios of the three PrPSc protein bands were compared. For ME7-infected C57BL/6J mice, the strongest PrPSc signal was observed in the mono-glycosylated band. However, for ME7-infected TgSShpPrP mice, the strongest PrPSc signal was observed in the di-glycosylated band (Fig. 2A). Similar glycoform patterns were observed from the first to the third passage (Fig. 2B). There was no difference between the electrophoretic mobilities of C57BL/6J-ME7 and TgSShpPrP-ME7 mice, although the C57BL/6J-ME7 pattern appeared to contain a greater amount of resistant PrPSc species after PNGase F treatment than that from TgSShpPrP-ME7 mice (Fig. 2C). The difference in glycoform patterns in the two mouse strains suggests that interspecies transmission of the infectious agent can result in a different glycoform characteristic in the new host.
Fig. 2
Abnormal PrP detection in ME7-infected C57BL/6J and TgSShpPrP mice. (A) Western blot analysis for PrPc and PrPSc in ME7-infected mice at 2nd passage. 20 μg of total protein were loaded into each well. For PrPSc detection, the brain homogenates were digested with 7 µg/mL of PK. PrPc and PrPSc were detected using mAb 7A12. Relative densities of un-, mono-, and di-glycosylated forms of PrPSc obtained from ME7-infected mice obtained using image J analyzer. (B) Western blot analysis for PrPc and PrPSc in ME7-infected mice at 1st, 2nd, and 3rd passages. Total protein (20 μg) was loaded into each well. In order to detect PrPSc, the brain homogenates were digested with 7 µg/mL of PK. The PrPc and PrPSc were detected using mAb 7A12. (C) To confirm the electrophoretic mobility of PrPSc from ME7-infected C57BL/6J and TgSShpPrP mice, brain homogenates were treated with PK and PNGase F prior to western blotting with mAb 7A12.
PrP, prion protein; TgSShpPrP, transgenic mouse line carrying the Suffolk sheep PrP gene; PrPc, cellular PrP; PrPSc, scrapie PrP; PK, proteinase K; PNGase F, peptide N-glycosidase F.
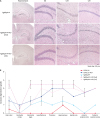
Spongiform degeneration in the hippocampal region of ME7-infected TgSShpPrP mice
Spongiform degeneration is known to be one of the hallmarks of neurodegeneration in prion disease [8]. To examine the extent of spongiform degeneration in ME7-infected TgSShpPrP brain tissue, using immunohistochemistry staining, hippocampal regions of control and infected mice brains were subjected to hematoxylin and eosin (H&E) staining. We observed an increase in spongiform degeneration in the dentate gyrus, cornu ammonis 3 (CA3) and CA1 hippocampal regions of infected mouse brains compared to that in the control mice brains (Fig. 3A). In prion diseases, the brain region profile is influenced by the prion's ‘strain properties,’ which indicate the invasion route to the brain [8]. We analyzed the histopathological alteration induced by the ME7 scrapie strain in the whole brain regions of TgSShpPrP mice. Nine regions of non-inoculated and ME7-inoculated TgSShpPrP mice brains were scored. The lesion scores of the ME7-inoculated TgSShpPrP mice were higher at both 2nd and 3rd passages than those of non-inoculated mice in most brain areas, with the exception of the cerebellar cortex (Fig. 3B).
Fig. 3
H&E staining and severity of neurodegeneration associated vacuolation in TgSShpPrP-ME7 infected brains. (A) H&E staining of age-matched control TgSShpPrP and terminally ill ME7-infected mice showing the dentate gyrus, CA3, and CA1 regions of the hippocampus. The hippocampus was viewed at a low magnification (4×); all other regions were viewed at a higher magnification (40×). (B) Mean vacuolation scores and standard errors of the means from brains of ME7-infected C57BL/6J and TgSShpPrP mice and age-matched control mice.
PrP, prion protein; H&E, hematoxylin and eosin; TgSShpPrP, transgenic mouse line carrying the Suffolk sheep PrP gene; DG, dentate gyrus; CA, cornu ammonis.
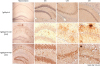
PrPSc detection in the hippocampal region of ME7-infected TgSShpPrP mice
To determine the localization and distribution of PrPSc in ME7-infected TgSShpPrP mice, aggregation and tissue deposition of PrPSc were observed in the area of the hippocampus. The most conspicuous feature of the PrPSc staining in ME7-infected TgSShpPrP brain was the heavy deposits of PrP plaques, which were mainly located in the dentate gyrus, CA3, and CA1 hippocampal regions (Fig. 4).
Fig. 4
PrPSc Immunostaining in the hippocampus of ME7-infected TgSShpPrP mice. PrPSc deposition in the DG, CA3, and CA1 hippocampal regions of age-matched control mice and ME7-infected mice (2nd and 3rd passages) were evaluated by using 3F10 anti-prion antibody. Plaques of an amyloid nature and plaque-like deposits (aggregated PrPSc) were observed in the ME7-infected brain regions. The hippocampus was viewed at a low magnification (4×); all other regions were viewed at higher magnification (40×).
PrP, prion protein; PrPSc, scrapie PrP; TgSShpPrP, transgenic mouse line carrying the Suffolk sheep PrP gene; DG, dentate gyrus; CA, cornu ammonis.
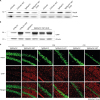
Extent of neuronal loss and activation of reactive astrocytes in ME7-infected C57BL/6J and TgSShpPrP mice
Notable features of prion diseases include activated astrogliosis and microglial and neuronal loss, and these neuropathological features are associated with the accumulation of a conformationally altered PrPSc isoform. We observed decreases in the expression level of NeuN in ME7-infected C57BL/6J and TgSShpPrP mice when compared to that of age-matched non-infected mice (Fig. 5A and C). However, ME7-infected mice brain showed increased expression of GFAP relative to that in non-infected mice (Fig. 5B and C). Immunohistochemical staining results indicated that GFAP immunoreactivity was higher in ME7-infected brains.
Fig. 5
Loss of neurons and activation of reactive astrocytes in ME7-infected C57BL/6J and TgSShpPrP mice at 2nd passage. Presence of GFAP indicating astrogliosis and loss of neurons were examined by immunoblotting (A, B) and immunostaining (C) using anti-GFAP and anti-NeuN in C57BL/6J and TgSShpPrP age-matched control mice and mice inoculated with ME7 strain (2nd passage). Both western blot and immunohistochemical results revealed increased expression of GFAP protein and decreased expression level of NeuN in ME7-infected mice relative to the expressions in the control mice.
PrP, prion protein; TgSShpPrP, transgenic mouse line carrying the Suffolk sheep PrP gene; GFAP, glial fibrillary acidic protein; DG, dentate gyrus; CA, cornu ammonis.
DISCUSSION
Scrapie can be experimentally induced in one species by transmission from another species. Interspecies transmission, when successful, results in prolonged incubation periods in the new host. This phenomenon, also known as a transmissibility barrier, has been shown to be abrogated with adaptation of the strain to the new host after 2–3 passages [9101112]. In this study, the transmissibility of a long-time mouse-adapted ME7 scrapie strain to TgSShpPrP mice was examined. We observed reductions in incubation time from 227.0 ± 6.35 days at first passage to 185.3 ± 5.41 days at second passage and 132 ± 1.55 days at 3rd passage. Consistent with our result is the report that cross-species transmission leads to an inefficient disease process resulting in long incubation time at first passage but becomes shortened after the prion becomes adapted to the new host. This is probably because there is increased efficiency of propagation of the donor PrPSc and the experimentally induced scrapie at second passage as it becomes more adapted to the environment of the recipient host [131415].
The glycosylation profile is one of the biochemical features used to discriminate scrapie strains. Results from our study indicate a change in glycoform patterns; the PrPSc from ME7-infected TgSShpPrP mice brains were predominantly di-glycosylated in contrast to the mono-glycosylated band pattern observed in ME7-infected C57BL/6J mice brains. Similar to our results are those reported by Shi et al. [1]. Similar glycosylation patterns observed at the third passage suggests that the glycosylation profile formed during interspecies transmission can be maintained over serial passaging in the same host [16]. This may also support the suggestion that the brain microenvironment of a new host can have a profound effect on PrPSc properties following interspecies transmission.
To determine the neuropathological changes induced by the interspecies transmission in this study, H&E staining was performed on formalin-fixed tissues of ME-7-infected TgSShpPrP and control mice. The dentate gyrus, CA3, and CA1 hippocampal regions were examined for spongiform degeneration. Our results showed that sponge-like lesions were more pronounced in the CA1 regions of the terminally ill ME7-infected TgSShpPrP mice examined. Numerous oval shaped vacuoles were observed in the fields of view, most particularly in the CA1 hippocampal region of the infected brains. Another criterion employed for determining strain peculiarity is the vacuolar lesion profile. Severe vacuolation was observed in the hippocampal, brain stem, and substantia nigra regions of infected mouse brains. Other grey matter areas such as hypothalamus, thalamus, and septal area also showed marked vacuolation but there was less conspicuous vacuolation in the cerebellar cortex, which is in agreement with results from a similar study report by Cunningham et al. [17]. Reports from other experimental studies have revealed that the pattern of PrPSc deposition across brains of infected mice is agent strain dependent [181920]. In this study, we detected PrPSc accumulation and the formation of plaques, and PrPSc aggregation accompanied by plaques was more prominent in the CA1 hippocampal region. Sisó et al. [21] reported that amyloid plaques are abundant and characteristics of murine 87V infection, but they inconsistently occur in ME7 and 22A infections, whereas vacuolar plaques are sometimes observed in ME7 infections of VRQ/VRQ Cheviot and ARQ/ARQ Suffolk sheep. In addition, Cunningham et al. [17] reported that ME7-infected brains showed a greater amount of the PrPSc punctuate labeling pattern with some small plaque depositions in both the hippocampal and thalamic brain regions. A strong counterstain is a well-known marker of sick cells about to die via an apoptotic pathway [2223]. Our results showed a decreased population of neuronal cells and a significant high accumulation of hematoxylin-counterstaining. Also, there was a decrease in the expression level of a neuronal marker used to measure the extent of neuronal loss between infected and control mice. Hippocampal CA1 pyramidal neuronal loss has been reported to be significant in ME7- and 79A-infected mice [17242526]. Most likely, the severe aggregation of PrPSc in CA1 in this study resulted from the death of CA1 neurons. The increase in the population of reactive astrocytes was well pronounced in brain tissues of the ME7-infected mice. Furthermore, GFAP upregulation was marked in the dentate gyrus of the brain, which is consistent with similar findings reported by Hilton et al. [25]. They demonstrated that the hippocampus and dentate gyrus are sites of GFAP upregulation in ME7- and 79A-infected animals.
Differences in pathological features such as the patterns of histological lesions and PrPSc accumulations in scrapie-infected mice have been reported to represent the total interaction between the host (genotype, age, sex, and breed) and the agent strain (passage history, dose, route of inoculation, donor genotype, and organ used for preparing inoculum) [62728]. In addition, it has been suggested that some disease phenotype features might be strain-specific and propagated in the recipient host [21].
In conclusion, interspecies transmission of a long-term mouse-adapted ME7 scrapie strain was successful in transgenic mice expressing ovine PrP and the infection was accompanied by biophysical and neuropathological changes in the recipient host. The differences in survival time, biophysical (glycoform change), and neuropathological features (plaque formation) reported in this study are most likely species dependent rather than strain encoded.
 XML Download
XML Download