

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Porcine pleuropneumonia is a highly contagious respiratory disease that causes massive economic losses in the global pork industry [123]. Actinobacillus pleuropneumoniae is known to be the major causative pathogen of this disease [12]. It has been reported that there are 15 serotypes of A. pleuropneumoniae, showing susceptibility to pigs of all ages [456]. Pigs recovered from acute infection and chronically infected pigs can be carrier animals that shed the pathogen continuously without clinical symptoms. Thus, it is difficult to control and eradicate this disease in infected herds [47].
Apx toxins, which belong to the pore-forming repeat-in-toxin (RTX) family of A. pleuropneumoniae, are known to play an important role in the pathogenesis of A. pleuropneumoniae as major virulence factors [48]. The importance of Apx toxins in protective immunity against A. pleuropneumoniae infection has also been reported in many studies [49101112]. There are four different Apx toxins, ApxI, ApxII, ApxIII, and ApxIV, each of which are encoded by the apx operon. The apx operon includes four genes, the activator gene of apxC, the pretoxin structural gene of apxA, and the secretion-apparatus-encoding genes of apxB and apxD [41112]. Among these toxins, ApxI is strongly hemolytic and cytotoxic, while ApxII is weakly hemolytic and moderately cytotoxic, and ApxIII is nonhemolytic and strongly cytotoxic. The toxins of ApxI, ApxII, and ApxIII, which are secreted by different serotypes in various combinations, mainly determine the virulence of the 15 serotypes of A. pleuropneumoniae [41011]. The ApxIV toxin is produced by all serotypes of A. pleuropneumoniae in infected pigs, but not under in vitro culture conditions; therefore, antibody response to ApxIV toxin has been used as a marker for infected pigs [410111314].
In an effort to control the disease, vaccination has been actively conducted in the swine industry around the world. Currently available vaccines can be classified based on the antigens such as bacterins (killed whole bacteria), purified toxoid vaccines, and mixed vaccines (killed whole bacteria and purified toxins) [101415]. Most of these A. pleuropneumoniae vaccines are primarily based on immunization against Apx toxins, which could be achieved effectively using subunit vaccines with recombinant proteins of ApxI, ApxII, and ApxIII [1015]. It has also been reported that subunit vaccines using recombinant Apx toxins showed safety and cross-protection against different serotypes of A. pleuropneumoniae [16]. Therefore, subunit vaccines are being applied worldwide and the development of new subunit vaccines is actively being conducted [1617181920].
It is important to measure immune responses to each Apx toxin separately when evaluating vaccine efficiency during the development of subunit vaccines containing the recombinant toxin antigens of ApxI, ApxII, and ApxIII [4721]. Immune responses to A. pleuropneumoniae could be detected using various diagnostic methods such as radioimmunoassay, hemolysin neutralization, the microplate agglutination test, and complement fixation [52223]. However, these approaches are not suitable for evaluation of the immune responses to each Apx toxin. Enzyme-linked immunosorbent assay (ELISA) using purified recombinant protein antigens is more sensitive than other methods and is able to determine antibody titers against each antigen [5242526]. Thus, ELISA methods have an advantage in evaluating vaccine efficiency for the development of subunit vaccines of A. pleuropneumoniae [3192026]. However, currently available commercial ELISA methods are difficult to apply to the evaluation of vaccine efficiency because they have been developed only to diagnose serotypes of A. pleuropneumoniae, while they cannot measure antibody titers against ApxI, ApxII, and ApxIII, respectively [112425]. Moreover, antibodies against Apx toxins have been reported to have cross-reactivity to each other, making the determination of levels of immune responses to each toxin more difficult [21272829]. Therefore, the development of ELISA methods that can detect specific antibodies against each toxin should take precedence for the development of A. pleuropneumoniae subunit vaccines.
In this study, we developed ELISA methods that can evaluate specific antibodies against each toxin of ApxI, ApxII, and ApxIII. Specific peptide regions in each toxin were identified and candidate partial Apx toxins for ELISA were purified. The ELISA conditions using the recombinant partial toxins were then established. Using the sera of guinea pigs vaccinated with each toxin of ApxI, ApxII, and ApxIII of the commercial vaccine, the identified ELISA conditions were assessed to determine if they could be used in vaccine development as a tool for vaccine efficiency evaluation based on the quantification of ApxI, ApxII, and ApxIII-specific antibody responses.
MATERIALS AND METHODS
Bacterial strains and culture conditions
A. pleuropneumoniae serotype 2, isolated in Korea, was used for the genes of apxIIA and apxIIIA [12]. The apxIA gene was obtained from A. pleuropneumoniae serotype 5 isolated from a Korean pig with pleuropneumonia. The bacteria were cultured in Luria-Bertani (LB) broth (Difco, USA) with 0.01% β-nicotinamide adenine dinucleotide for 24 h at 37°C.
Homology analysis of apxIA, apxIIA, and apxIIIA
Protein sequence information regarding apxIA (AF363361), apxIIA (AF363362), and apxIIIA (AF363363) was retrieved from the National Center for Biotechnology Information database. Homology and conserved domains were investigated using multiple sequence alignment of the ClustalW2 program (http://www.ebi.ac.uk/Tools/clustalw) to identify specific gene regions among toxins.
Identification and cloning of specific genes
The gene sequence of each toxin was divided into four regions, the N-terminal (Nt), middle part 1 (M1), middle part 2 (M2), and C-terminal (Ct), for investigation of specific gene regions. Specific gene regions showing low levels of homology were determined based on the results of alignment of the four gene regions of each toxin. The specific gene regions identified for each toxin (partial Apx toxins) were amplified by polymerase chain reaction (PCR) using the primers described in Table 1. Total genomic DNA extracted from A. pleuropneumoniae serotype 2 and 5 was used for PCR templates (GenElute; Sigma-Aldrich, USA). The PCR products were electrophoresed in 1.0% agarose gel and then purified with a gel extraction kit (QIAquick; Qiagen, Germany). Amplified gene regions for each toxin were cloned with the Chapion pET100 vector (Invitrogen, USA) into Top10 chemical-component Escherichia coli cells (Invitrogen) according to the manufacturer's protocols. To confirm gene insertion, DNA sequencing was conducted using an automated DNA sequencer (ABI377L; Applied Biosystems, USA). Following confirmation of gene insertion by DNA sequencing, the cloned partial Apx toxins were transformed into component E. coli M15 cells for expression.
Table 1
Primers for recombinant partial toxins of ApxIA, ApxIIA, and ApxIIIA

![]()
Expression and purification of the recombinant proteins
E. coli M15 cells were grown in LB broth with ampicillin (100 µg/mL) at 37°C. When the culture reached an optical density (OD) of 0.6 nm, isopropyl β-D-1-thiogalactopyranoside (1 mM; Duchefa Biochemie, the Netherlands) was added and then cultured continuously for 4 h. The harvested cells were subsequently re-suspended in lysis buffer (20 mM Tris-hydrogen chloride, 500 mM sodium chloride, 8 M urea, 40 mM imidazole, pH 7.0). Nickel-nitrilotriacetic acid chelate affinity chromatography (GE Healthcare, UK) was conducted based on the manufacturer's protocols, after which the bound proteins were eluted with elution buffer (20 mM Tris-hydrogen chloride, 500 mM sodium chloride, 8 M urea, 500 mM imidazole, pH 7.0). The concentration of the purified recombinant proteins was then determined using a BCA Protein Assay kit (Pierce, USA)
Sodium dodecylsulfate-polyacrylamide gel electrophoresis and Western blot analysis
The purified partial-Apx toxins were separated by 12% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and stained with Coomassie blue R-250. For Western blot analysis, the separated proteins from the SDS-PAGE were electrotransferred onto an iBlot Gel Transfer Stacks nitrocellulose membrane (Invitrogen), which was then immunoblotted with sera (1:1,000) of guinea pigs vaccinated with subunit vaccines containing ApxI, ApxII, and ApxIII toxoids, respectively. An alkaline phosphatase-conjugated rabbit anti-pig IgG (H+L) at 1:2,000 dilution was used as the secondary antibody and the specific reaction was detected using an AP-conjugated substrate kit (BioRad, USA). The predicted size of the purified partial-Apx toxins matched the results of the SDS-PAGE and Western blot analyses based on amino acids constitution.
Guinea pig vaccination with ApxI, ApxII, and ApxIII toxoids
The full-length ApxIA, ApxIIA, and ApxIIIA are recombinant proteins and major components of the A. pleuropneumoniae vaccine newly developed by Choong Ang Vaccine Laboratories Company (Daejeon, Korea). Guinea pig vaccination was carried out three times with these full-length Apx toxoids by intramuscular injection of 200 µg of each toxin at two-week intervals, respectively. Sampling of sera was performed two days after the last administration. All experiments using guinea pigs were conducted in accordance with the procedures permitted by the Institutional Animal Care and Use Committees of Choong Ang Vaccine Laboratories Company (No. 160129-01).
Development enzyme-linked immunosorbent assays for ApxIa, ApxIIa, and ApxIIIa-specific detection
The assays were developed through checkerboard titration of 96-well microplates utilizing representative positive and negative sera. Sensitization of partial recombinant ApxIA, ApxIIA, and ApxIIIA antigens was performed using a two-fold serial dilution of each antigen at levels ranging from 3.125 ng/well to 200 ng/well. Antigens of partial-recombinant Apx toxins were coated onto 96-well microplates with 50 mM coating buffer (15 mM sodium carbonate and 35 mM sodium bicarbonate, pH 9.6), then incubated at 4°C overnight. Next, microplates were washed with a solution of phosphate-buffered saline containing 0.05% Tween 20 (PBST), after which they were blocked at room temperature for 2 h using 3% bovine serum albumin fraction V (BSA; Fisher Scientific, USA) in PBST. Sera from vaccinated guinea pigs with each full-length Apx toxin and serum from a nonvaccinated guinea pig were used as positive and negative controls, respectively. The microplates were incubated at room temperature for 2 h with 100 µL of two-fold serial diluted sera, after which they were incubated with 100 µL of 1:10,000 diluted horseradish peroxidase-conjugated goat anti-guinea pig IgG (H+L) (BioRad) at room temperature for 1 h. Sera and antibody used in ELISA were diluted with 1% BSA in PBST. Color development was performed with 3,3′,5,5′-tetramethylbenzidine liquid substrate (Sigma-Aldrich) at room temperature for 10 min. The color development was stopped with 1 N sulfuric acid, after which the microplates were read at 450 nm. Based on the optimal ELISA conditions confirmed by checkerboard titration, ApxI, ApxII, and ApxIII-specific ELISA conditions were established under the same conditions of the checkerboard titration; however, 5% (for blocking) and 3% (for dilution of sera and antibody) skim milk in PBST were used.
RESULTS
Homology among apxIA, apxIIA, and apxIIIA
All of the Apx toxins showed > 40% homology with each other (Table 2). The results of homology analysis between regions within each toxin (Nt, M1, M2, and Ct) are presented in Table 3. The M2 region was considered to be inadequate for the specific gene region based on high levels of homology. Based on the low percent identity observed upon homology analysis, specific gene regions were determined for ELISA antigen candidate peptides as follows: Nt and Ct regions in ApxIA; Nt, M2, and Ct regions in ApxIIA; Nt, M2, and Ct regions in ApxIIIA. The determined sequence regions are shown in yellow in Fig. 1.
Table 2
Homology among ApxIA, ApxIIA, and ApxIIIA

| Proteins and accession No. | Percent identity (%) | |||
|---|---|---|---|---|
| ApxIA | ApxIIA | ApxIIIA | ||
| ApxIA (1022aa) | ||||
| AF363361 | - | 44.0 | 50.9 | |
| ApxIIA (956aa) | ||||
| AF363362 | 44.0 | - | 44.6 | |
| ApxIIIA (1052aa) | ||||
| AF363363 | 50.9 | 44.6 | - | |
![]()
Table 3
Homology among the Nt, M1, M2, and Ct regions within Apx toxins

![]()

Production of the recombinant partial toxins of ApxIA, ApxIIA, and ApxIIIA
Based on the results of homology analysis, all selected specific gene regions except ApxIIA Ct were amplified using the primers shown in Table 1 and then cloned for production of recombinant proteins. As shown in Fig. 2, the molecular masses of the recombinant proteins were in concordance with the predicted molecular weights (ApxIA Nt, 19.3 kDa; ApxIA Ct, 24.2 kDa; ApxIIA Nt, 19.1 kDa; ApxIIA M2, 21.3 kDa; ApxIIIA Nt, 20.1 kDa; ApxIIIA M2, 22 kDa; ApxIIIA Ct, 25.9 kDa). Upon Western blot analysis, the recombinant proteins were found to be responsive to the positive control of guinea pig sera, with cross reactions observed between the purified partial toxins upon homology analysis as expected.
 | Fig. 2Confirmation of purification and antigenicity or recombinant partial Apx toxins by SDS-PAGE and Western blot. Upper and lower panels represent SDS-PAGE and Western blot, respectively. Western blot was conducted using positive guinea pig serum of ApxIA (A), ApxIIA (B), and ApxIIIA (C), respectively.SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrophoresis.
|
Development of ApxIA, ApxIIA, and ApxIIIA-specific enzyme-linked immunosorbent assays
Checkerboard titration was performed to establish the optimal ELISA conditions making a clear distinction between positive and negative sera. The optimal concentrations of recombinant partial toxins used to coat the microplates were as follows: 100 ng for ApxIA Nt, 200 ng for ApxIA Ct, 200 ng for ApxIIA Nt, 50 ng for ApxIIA M2, 200 ng for ApxIIIA Nt, 200 ng for ApxIIIA M2, and 100 ng for ApxIIIA Ct (Supplementary Figs. 1, 2, 3). To establish ELISA conditions for measuring ApxIA, ApxII, and ApxIIIA-specific antibody levels using the recombinant partial toxins, purified proteins of ApxIA Ct, ApxIIA Nt, and ApxIIIA Nt were selected as ELISA antigens considering the cross-reactivity (Fig. 3). Based on the checkerboard results and selected antigens, a 1:320 dilution ratio of guinea pig sera was found to show the most precise results. As shown in Fig. 3, ApxIA Ct (200 ng/well) had higher reactivity with ApxIA positive guinea pig sera than ApxIIA Nt (200 ng/well) and ApxIIIA Nt (200 ng/well). ApxIIA Nt and ApxIIIA Nt also showed greater responses to each positive serum than the other two antigens.
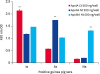 | Fig. 3The ELISA results for each Apx toxin (ApxIA, ApxIIA, and ApxIIIA)-positive guinea pig sera under the optimal conditions. The determined antigen concentrations (ng/well) and serum dilution rate (1:320) are described in the graph. Each Apx toxin-positive guinea pig serum could be identified by ELISA using ApxIA Ct, ApxIIA Nt, and ApxIIIA Nt.ELISA, enzyme-linked immunosorbent assay; Nt, N-terminal; Ct, C-terminal; **, p < 0.01.
|
DISCUSSION
Vaccination appears to be the most effective way to control A. pleuropneumoniae infection and has therefore been adapted in many countries around the world. It has also been reported that subunit vaccines against A. pleuropneumoniae using recombinant Apx toxins showed safety and cross-protection against different serotypes [16]. For this reason, many currently available vaccines use the recombinant Apx toxins as their key antigens [1015] and new subunit vaccines are actively being developed [1617181920]. Therefore, the measurement of immune responses to purified Apx toxin antigens has been an important milestone in vaccine development as a tool for vaccine efficiency evaluation [4721].
ELISAs are widely used to investigate immunization levels induced by A. pleuropneumoniae vaccines. However, most commercial ELISAs have been developed for serotype determination of A. pleuropneumoniae and cannot be easily applied to measure Apx-specific immune responses when evaluating the efficiency of Apx toxins-based subunit vaccines [112425]. In addition, experimental ELISAs reported in previous studies are also still difficult to differentiate levels of immunization against respective Apx toxin antigens [32629]. This problem is believed to be due to cross-reactivity among ApxI, ApxII, and ApxIII as a result of their high homology [21272829], which was also observed in homology analysis conducted in the present study (Table 2). By excluding antigen regions showing high homology among Apx toxins, therefore, we attempted to identify specific antigen regions to purify ELISA antigens for differential serological detection. To accomplish this, Apx toxin amino sequences were divided into regions of Nt, M1, M2, and Ct, and specific regions of Apx toxins were then cloned to obtain purified recombinant Apx toxin proteins. Nevertheless, positive guinea pig sera to each toxin of ApxI, ApxII, and ApxIII were reacted with all of the partial Apx toxins upon Western blot analysis. These nonspecific reactivities were thought to be attributed to the high levels of homology among partial proteins, as was observed during homology analysis (Table 3). Although specific antigen-antibody reactions were not observed upon Western blot analysis, the antigenicity of purified partial Apx toxins was confirmed.
To establish the optimal ELISA conditions for each recombinant partial toxin, checkerboard titration was conducted using guinea pig sera positive to each toxin of ApxI, ApxII, and ApxIII. Considering the need for ELISA to serve as a simple and convenient diagnostic tool, we attempted to set serum dilution conditions to the same ratio. Antigen coating conditions were determined to be the concentration at which the changes in the OD value of 450 nm were most prominent according to the serum dilution. Optimal ELISA conditions were attempted to be set to give an OD value of about 1.5 for each positive serum at the determined serum dilution and antigen concentration.
The results of Western blot and ELISA checkerboard titration suggested that the recombinant partial Apx toxins of ApxIA Nt, ApxIA Ct, ApxIIA Nt, ApxIIA M2, ApxIIIA Nt, and ApxIIIA Ct had a high possibility of being useful as ELISA antigens. The ApxIIIA M2 partial toxin was ruled out based on its low antigenicity to ApxIIIA positive guinea pig serum in the ELISA checkerboard titration. Based on the optimal ELISA conditions, an antigen combination of ApxIA Ct, ApxIIA Nt, and ApxIIIA Nt was determined by additional ELISA experiments (Fig. 3). In the additional ELISA experiments, all procedures were conducted using the optimal ELISA conditions; however, skim milk solution for blocking (5% in PBST) and antibody diluent (3% in PBST) were used. The skim milk solution was adapted to further reduce nonspecific reactions. As shown in Fig. 3, each of the antigens (ApxIA Ct, ApxIIA Nt, and ApxIIIA Nt) showed a higher OD value in response to each positive serum than the other two antigens. As reported in previous studies, cross-reactivity was also observed in the ELISA using ApxIA Ct and ApxIIA Nt antigens, but they could be distinguished based on the difference in OD value. In addition, ELISA using ApxIII Nt antigen showed excellent reactivity to its positive guinea pig serum when compared to the other two positive control guinea pig sera (Fig. 3).
The results of this study confirmed the high homology among toxins of ApxIA, ApxIIA, and ApxIIIA, which has been considered to be an important factor in the cross-reactivity of antibodies to each toxin. Due to this cross-reactivity, it has been difficult to measure and differentiate antibody levels to each of the three toxins. In this study, the recombinant partial Apx toxins were purified to reduce the interference of homology observed among the three toxins, and the produced partial Apx toxins were used to develop an ELISA method capable of measuring the level of each toxin-specific antibody. This Apx toxin-specific ELISA will be useful as an important tool for evaluation of vaccine efficiency by measuring specific antibody responses to each toxin during the development of subunit vaccines against A. pleuropneumoniae.
 XML Download
XML Download