

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Breast cancer can be classified into distinct subtypes based on its intrinsic molecular characterizations [123]. Breast cancer cell lines can also be grouped according to the intrinsic molecular subtype, which exerts different therapeutic responsiveness to chemotherapy [456]. Basal subtype breast cancers exhibit clinical characteristics of metastasis, invasiveness, high grade, and poor prognosis, and have been found to overlap with the triple-negative histological phenotype [78910]. However, only a few effective therapies exist for the patients with basal subtype breast cancer.
Pokemon (also known as FBI, ZBTB7A, LRF, and OCZF) is a member of the POZ/BTB and Krüppel (POK) family of transcription factors [11]. POK transcription factors are critical developmental regulators and have been directly implicated in human cancer. Over-expression of Pokemon has been reported in different types of human cancers of various origins, including non-small cell lung cancer, breast cancer, and ovarian cancer [12131415]. Pokemon was originally confirmed to function as a proto-oncoprotein by repressing the transcription of tumor suppressor gene alternate open reading frame (ARF) [16]. The expression of Pokemon in immature mouse T and B lymphoid lineages was found to cause aggressive lymphomas [17]. However, Pokemon inactivation profoundly promotes the progression of PTEN-loss driven prostate tumorigenesis by activating SRY-box 9 (SOX9)-dependent oncogenic pathways and promotes proliferation, survival, and invasion of cancer cells [18]. Recently, it was discovered that Pokemon directly represses the expression of key glycolytic genes, the loss of which led to the induction of glycolysis promoting tumor progression [19]. These findings argue against the proto-oncogenic role of Pokemon in human cancer, identifying Pokemon as a context-dependent cancer gene.
Cytokine transforming growth factor β (TGFβ) plays complex roles in carcinogenesis, being a tumor suppressor in the early stages and a tumor promoter in later stages of carcinogenesis [2021]. Smad4 is a central mediator of TGFβ intracellular signaling and it plays an important role in both tumor suppression and progression of breast cancer cells [2223]. Smad4 facilitates the formation of osteolytic bone metastasis and induction of IL-11, a gene implicated in bone metastasis [23]. However, Pokemon could function as a partner of Smad4 and play a negative role in the TGFβ pathway by interacting with Smad4 in HaCaT cells [24]. In a previous study [24], protein interaction between Pokemon and Smad4 was verified by the yeast two-hybrid screen, co-immunoprecipitation, and GST-pull down, among others. Pokemon decreased the recruitment of Smad4 co-activator p300/CBP and recruited histone deacetylase 1 (HDAC1) into Smad4-Pokemon complexes to inhibit the TGFβ pathway. However, Pokemon did not inhibit Smad4 DNA binding activity. As specificity protein 1 (SP1) has been found to be an important potentiator in the TGFβ/Smad4 signaling pathway [2526], in this study, we attempted to elucidate whether SP1 plays a role in the interaction between Pokemon and Smad4.
We have previously demonstrated the role of Pokemon in the progression of breast cancer via modulation of survivin expression [14]. In the present study, we sought to determine the effect of Pokemon on the proliferation ability of breast cancer cells and identify its role in Smad4 regulation during breast cancer progression.
METHODS
Cell lines
MDA-MB-231, T47D, and MCF-10A cell lines were purchased from the Chinese Academy of Sciences Cell Bank (Shanghai, China). The cells were cultured in Dulbecco's modified Eagle medium (Invitrogen, Waltham, USA) supplemented with 10% fetal bovine serum (Gibco, New York, USA). All of the cell lines were cultured in an atmosphere containing 5% CO2 at 37°C.
Plasmids and small interfering RNA
We constructed a Pokemon overexpression vector according to the previous method [27], with minor modifications. Briefly, full-length human Pokemon was obtained by standard polymerase chain reaction (PCR) amplification from MCF7 cell cDNA and was cloned into the green fluorescence protein (GFP)-linked pcDNA3.1 plasmid. Pokemon small interfering RNA (siRNA) and negative siRNA were bought from GenePharma (Shanghai, China).
Quantitative real-time PCR
Cells were lysed using Trizol reagent (Life Technology, Rockville, USA). The cell lysates were mixed with chloroform and divided into aqueous phase and organic phase. Total RNA was extracted from the supernatant. Reverse-transcription was completed using PrimeScript™ RT Reagent Kit (Takara, Dalian, China), which catalyzes RNA to cDNA. Polymerase chain reaction for cDNA was induced by SYBR Green PCR Master Mix (Takara). Final detection for transcriptional expression was performed using a 7500 real-time PCR System (Applied Biosystems, Foster City, USA). The mRNA expression levels were normalized to the house-keeping gene GAPDH and were calculated using the 2−ΔΔCt method.
Gene microarray analysis
Gene microarray was performed as previously described [28]. Briefly, after isolation of total RNA and reverse transcription, cDNA was then subjected to gene expression profiling using the Affymetrix Human Gene 1.0T arrays (Genechem, Shanghai, China). Data were obtained by Genspring 7.0, Expression Console version 1.1.1, and DNA-chip Analyzer 2008 (dChip). The gene expression data were analyzed using the SBC Analysis System (Genechem). The retrieved data showing a fold-change ≥ 1.5 were filtered out. These genes were then functionally classified based on Gene Ontology Database, Affymetrix Database, and DAVID 6.7 Functional Annotation Database.
EdU incorporation assay
Cells were transfected with Pokemon siRNA and treated with TGFβ1 or not for 48 hours. EdU (5-ethynyl-2′-deoxyuridine, 50 µM; Cell-Light™ EdU Apollo®488 In Vitro Imaging Kit; RiboBio, Guangzhou, China) was added to further culture for 2 hours. According to the manufacturer's protocol, the EdU-containing medium was removed and 4% paraformaldehyde was used to fix cells at room temperature for 30 minutes. After removing the paraformaldehyde, lysine (2 mg/mL in deionized water) was added under shaking for 5 minutes, followed by 2 phosphate-buffered saline (PBS) washes. Then, Apollo 480 fluorescent azide reaction buffer was added and allowed to react for 30 minutes in darkness, before washing with 0.5% Triton X-100. After staining with Hoechst dye for 30 minutes, cells were washed with PBS and were finally stored in 100 µL PBS. Images were obtained using a fluorescence microscope. The percentage of EdU positive cells was calculated as follows: (EdU Incorporated Cells/Hoechst Stained Cells) × 100%.
MTS assay
Cells were treated with Pokemon siRNA for 24 hours, transferred into a 96-well plate with a density of 4 × 103 cells per well, and then treated with TGFβ1 or not for 48 hours. Then 20 µL of MTS reaction buffer was added per well, and cells were further cultured for 2 to 4 hours at 37°C. The optical density values represented the cell viability and were obtained using a microplate reader at a wavelength of 490 nm.
Colony formation assay
After transfection, cells were cultured in normal culture medium for 14 days. Cells were fixed with 4% paraformaldehyde for 30 minutes at room temperature and then washed twice with PBS. Wright-Giemsa Stain (Baso, Zhuhai, China) was used to stain cell colonies according to the protocol. Briefly, 10 drops of Giemsa solution A were added and allowed to stain for 3 minutes. This was then rapidly mixed with 20 drops of Giemsa solution B, followed by shaking of the cell plates for 5 minutes. The stain solution was then washed with flowing water and finally the plates were air dried. Colony number was counted by direct visual counting.
Western blotting
Total proteins were obtained from the supernatant lysis buffer of breast cancer cells. A BCA protein assay kit (Beyotime, Shanghai, China) was used to measure the protein concentrations according to the manufacturer's instruction. Proteins with different molecular weights were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, USA). Membranes were incubated in 5% non-fat milk for 1 hour at room temperature, then at 4°C with primary antibodies overnight, followed by 1 hour at room temperature with the corresponding secondary antibodies. Protein bands were shown by ECL Reagent (Beyotime) and were photographed using the Fluorchem E System (Cell Biosciences, Santa Clara, USA). The antibodies used for GFP-Pokemon, Smad4, and β-actin were obtained from CST (Boston, USA). The secondary antibodies were obtained from Proteintech Group (Chicago, USA).
Immunofluorescence analysis
Cells were collected and fixed with 75% ethanol at −20°C and were blocked with 2% bovine serum albumin in PBS for 1 hour at room temperature. Then, cells were incubated with primary rabbit anti-TGFβ (1:100, CST) or rabbit anti-Smad4 (1:100, CST) antibodies overnight at 4°C. After 3 washes with PBS, cells were incubated with secondary antibodies (anti-rabbit IgG conjugated with Alexa Fluor 594, 1:1,000, CST) for 1 hour. Cell nuclei were shown by DAPI (or 4′,6-diamidino-2-phenylindole) staining. Images were obtained using a laser scanning confocal microscope (LSM510; Carl Zeiss, Jena, Germany).
Co-immunoprecipitation assay
In order to obtain total protein, transfected cells were harvested and lysed using lysis buffer containing protease inhibitors. Co-immunoprecipitation assay was performed using a co-immunoprecipitation kit (Pierce, Rockford, USA) according to the manufacturer's instructions. Briefly, 3 mg of total protein for each treatment was used in this study, pre-cleared with sepharose resins. The supernatants were divided equally into 2 tubes and incubated into columns containing 1 µg immobilized anti-Pokemon or anti-SP1 antibody (CST). The immunocomplexes were covalently associated with sepharose resins. The resins were eluted 5 times and the co-immunoprecipitates were separated on an SDS-PAGE gel before transferring to a PVDF membrane for analysis with anti-Pokemon or anti-SP1 antibodies.
Luciferase reporter assay
MCF-7 or MDB-MA-231 cells were seeded into a 6-well plate (2 × 105 cells per well) and were incubated overnight. The cells were then co-transfected with either 0.3 µg/well of Smad4 firefly luciferase reporter plasmid constructs (pLuc366 or pLuc207) or the control pGL3-Basic vector (Promega, San Luis Obispo, USA), then respectively co-transfected with or without Pokemon- and SP1-expressing plasmids. The renilla luciferase plasmid was also co-transfected to correct for variations in transfection efficiency (50 ng/well). After incubating for 48 hours, the cells in each well were separately collected and the luciferase activity was measured using a microplate luminometer. Final data were calculated as the ratio of firefly luciferase activity versus renilla luciferase activity units (n = 3).
Electrophoretic mobility shift assay
Smad4 promoter probes (Probe 1 and Probe 2) were synthesized by Sangon Biotech (Shanghai, China). Probe 1: 5′-AGCGGGAGGCGGGGGCAGCC-3′ for the wild-type promoter (Probe); 5′-AGCAAAAGGCAAAAACAGCC-3′ for the mutant promoter (mProbe). Probe 2: 5′-AAAGCTGCGGGGGAAAAG-3′ for the wild-type promoter; 5′-AAAGCTGCAAAAAAAAAG-3′ for the mutant promoter. Probes were labeled with biotin. Nuclear extracts of MDB-MA-231 cells were prepared using a nuclear extraction kit (Pierce). In brief, an electrophoretic mobility shift assay (EMSA) was performed as follows: nuclear extracts (20 µg) and labeled probes (200 fM) were incubated for nuclear extract binding reaction; probes without nuclear extracts were used as the negative control; a combination of nuclear extracts, probes (Probe or mProbe), and unlabeled probe (com-Probe, 200×) were used for the competition reaction; a combination of nuclear extracts, probes, and IgG, anti-Pokemon or anti-SP1 antibodies were used for the supershift reaction. DNA-protein complexes were loaded onto a 6% native polyacrylamide gel. After electrophoresis, separated DNA was transferred to a nylon membrane before being cross-linked and photographed.
Statistical analysis
Analyses in this study were performed using the GraphPad Prism statistical software (GraphPad Software, La Jolla, USA). Experimental data were represented as the mean ± standard error of the mean. Statistical analysis was performed via one-way analysis of variance followed by Dunnett's test for multiple comparisons. The p-values less than 0.05 were considered statistically significant.
RESULTS
Pokemon deletion aggravates TGFβ-induced cell proliferation arrest in breast cancer cells
In order to verify whether Pokemon acts as a pro-oncoprotein or a tumor suppressor protein, and whether Pokemon promotes or inhibits cell proliferation arrest in breast cancer cells, cell proliferation ability-related experiments were applied (Figure 1). For alternative Pokemon downregulation or TGFβ treatment in MDB-MA-231 cells, DNA replication ability verified by EdU incorporation assay was significantly suppressed (Figure 1A and B). The cell proliferation rate, measured by MTS assay, was reduced (Figure 1C). Cell viability observed by optical photograph was reduced (Figure 1D and E) and cell colony numbers were less than control cells (Figure 1F and G). Co-treatment of Pokemon siRNA and TGFβ proteins presented contrasting results. This data highlights the key inhibition role of Pokemon in TGFβ1-induced cell proliferation arrest in breast cancer cells.
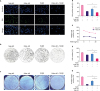 | Figure 1Aggravation of TGFβ-induced cell proliferation arrest after Pokemon deletion in breast cancer cells. After transiently transfected with Neg-siR, Poke-siR (100 nM) or treated with TGFβ1 (10 ng/mL) for 48 hours in MDA-MB-231 cells (A, B), the number of EdU positive cells (EdU+, green color), which represented DNA replication ability, was reduced than control group. (C-E) Alternative siRNA or TGFβ treatment for 48 hours, the viability and proliferation of breast cancer cells were significantly weakened. (F) The colony number of breast cancer cells was also decreased by the 2 treatments. In the above experiments, Pokemon deficiency could promote the growth arrest by TGFβ protein treatment (n = 4).TGFβ = transforming growth factor β; Neg-siR = negative siRNA; Poke-siR = Pokemon siRNA.
*p < 0.01, †p < 0.001.
|
Pokemon modulates TGFβ-Smad4 pathway in breast cancer cells
To elucidate the mechanism by which Pokemon is involved TGFβ-dependent cell proliferation arrest in breast cancer cells, we used gene microarray to screen for signaling related to Pokemon expression in MDB-MA-231 cells. Several signaling pathways, such as those of the metabolism of lipids and lipoproteins, hemostasis, and the adaptive immune system, were revealed to be responsive to Pokemon alteration (Figure 2A). We also found that “pathway in cancer”-associated genes, such as TGFβ, Smad4, TGFα, MYC, and BIRC3I, were repressed by ectopic Pokemon expression in MDB-MA-231 cells (Figure 2B). Ectopic Pokemon expression also inhibited the expression of Smad4, as detected by immunofluorescence (IF) assay and western blotting (Figure 2C-F).
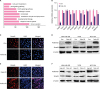 | Figure 2Negative connection between Pokemon and TGFβ-Smad4. (A) Up-regulation of Pokemon affected a series of cell signaling pathway, which was evaluated by the gene array analysis. (B) The mRNA levels of cell invasion-related genes were detected by quantitative real-time polymerase chain reaction after the transfection of Poke-OE or Con. (C) IF assay identified up- or down-regulation of Pokemon reduced or increased expression of Smad4. (D) Western blot results indicated transfection of over-expressed plasmid of Pokemon reduced protein levels of Smad4, and the Pokemon siRNA transfection showed the adverse effects (n = 4).TGFβ = transforming growth factor β; Poke-OE = Pokemon over-expression plasmid with GFP label; Con = negative plasmid; IF= immunofluorescence; MAPK= mitogen-activated protein kinase; DAPI = 4',6-diamidino-2-phenylindole.
*p < 0.05, †p < 0.01.
|
Pokemon regulates the expression of multiple proliferation-associated genes in the TGFβ-Smad4 pathway
We further determined the effect of Pokemon on the expression of proliferation-associated genes in the TGFβ pathway by using PCR gene expression array in breast cancer cells with Pokemon siRNA. This analysis identified multiple genes involved in TGFβ signaling-associated cellular proliferation regulated by Pokemon in breast cancer cells. The expression levels of 13 key genes associated with cellular proliferation during TGFβ signaling were analyzed. Pokemon siRNA was found to result in the downregulation of CYCLIND1, CDKN1A, GADD45B, and CDC6, and the upregulation of TGFβ1 in both MDB-MA-231 and T47D cells. TGFβ2, TGFβ3, CDKN1B, IGFBP1, ATF3, EMP1, PTK2, and ACVR1 showed distinct responses to the downregulation of Pokemon in MDB-MA-231 and T47D cells (Figure 3A and B). The levels of the genes above, with the exception of IGFBP1, were all downregulated after Pokemon siRNA transfection in MCF-10A cells (Figure 3C). Western blotting demonstrated that the protein levels of CYCLIND1 and CDKN1A also decreased after Pokemon knockdown in three breast cancer cell lines (Figure 3D). These results suggest that Pokemon regulates multiple proliferation associated genes in the TGFβ pathway and could thus be an essential modulator in TGFβ pathway-induced proliferation inhibition in breast cancer cells.
 | Figure 3Pokemon regulation on multiple proliferation associated genes. (A-C) Quantitative real-time polymerase chain reaction assay identified that Pokemon knockdown increased TGFβ1 gene level and reduced several proliferation-associated gene levels in MDA-MB-231, T47D and MCF-10A cells (n = 4). (D) Protein levels of CYCLIND1 and CDKN1A were reduced by Pokemon knockdown.TGFβ = transforming growth factor β; Neg-siR = negative siRNA; Poke-siR = Pokemon siRNA; CDKN1A = cyclin dependent kinase inhibitor 1A; CDKN1B = cyclin dependent kinase inhibitor 1B; GADD45B = growth arrest and DNA-damage-inducible, beta; IGFBP3 = insulin-like growth factor-binding protein 3; ATF3 = activating transcription factor 3; CDC6 = cell division cycle 6; EMP1 = epithelial membrane protein 1; PTK2 = protein tyrosine kinase 2; ACVR1 = activin A receptor, type I.
*p < 0.05, †p < 0.01, ‡p < 0.001.
|
Pokemon was bound with SP1 to indirectly suppress Smad4 promoter activity
To further identify the mechanism of Pokemon-induced Smad4 repression, we used a dual luciferase reporter system to assess the role of Pokemon in the transcription-regulation of Smad4. Two specific luciferase plasmids containing the Smad4 proximal promoter sequence, pLuc366 and pLuc207, were used to test Smad4 promoter activity. As a result, Pokemon was found to reduce Smad4 luciferase activity in MDB-MA-231 cells (Figure 4A). Furthermore, an EMSA was used to confirm whether the Pokemon protein bound to the Smad4 promoter sequences. Two specific Smad4 promoter probes (Probe 1 and Probe 2) with a GC box sequence were used to bind with the nucleus extraction obtained from MDB-MA-231 cells. Pokemon antibody was found to be unable to trigger the supershift of the protein DNA complex (Figure 4B), indicating that Pokemon has no direct protein-DNA interactions with the Smad4 promoter sequences. SP1, which was previously proved to regulate Smad4 promoter activity during invasion, was found to bind with Probe 1. Moreover, the ectopic expression of Pokemon was revealed to decrease the binding affinity of SP1 with Probe 1. The co-immunoprecipitation assay also showed the direct interaction between SP1 and Pokemon in MDB-MA-231 cells (Figure 4C), and a dual luciferase reporter assay verified that the SP1 enhanced-promoter activity of Smad4 was abrogated by Pokemon in MDB-MA-231 cells (Figure 4D). These results indicated that Pokemon functions as a negative modulator of Smad4 expression by interacting with SP1, leading to the separation of SP1 from the Smad4 promoter sequence, leading to decreased transcription activity of Smad4 in breast cancer cells.
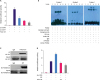 | Figure 4Indirect inhibition of Pokemon on Smad4 promoter activity through binding with SP1. (A) Dual luciferase reporter system showed that transfection of Poke-OE reduced Smad4 luciferase activity (pLuc366 and pLuc207, Smad4 expression plasmid vectors in MDB-MA-231 cells). (B) EMSA assay applied with 2 Smad4 probes proved Pokemon was not bound to promoter active regions of Smad4. (C) Co-immunoprecipitation identified that Pokemon was closely bound to SP1 protein. (D) SP1 promoted luciferase activity of Smad4, which was dose-depend weaken by Pokemon vectors (0.5 µg, 1 µg) in breast cancer cells (n = 3).SP1 = specificity protein 1; Poke-OE = Pokemon over-expressing vectors; IP = immunoprecipitation; IB = immunoblotting; Ab = antibody.
*p < 0.05, †p < 0.01.
|
Pokemon and Smad4 showed contrasting expression levels in different types of breast cancer
After confirming that Pokemon decreases the expression of Smad4 and blocks breast cancer cell growth in vitro, the correlation between the expression of these 2 proteins was identified in clinical samples. To further validate the correlation between Pokemon and Smad4 expression in a broad range of clinical samples, we analyzed public gene expression microarray datasets of breast cancer tissues from 2 The Cancer Genome Atlas (TCGA) datasets: Breast Invasive Carcinoma (TCGA, Cell 2015; http://www.cbioportal.org/study?id=brca_tcga_pub2015#summary) and Breast Invasive Carcinoma (TCGA, Nature 2012; http://www.cbioportal.org/study?id=brca_tcga_pub#summary). Consistent with our in vitro data, the Pokemon mRNA levels showed a significantly negative correlation with Smad4 mRNA levels in different subtypes of breast cancer cases across the two independent datasets. Pokemon showed the highest mRNA levels in the subtype of Luminal A and the lowest levels in the basal-like subtype (Figure 5A and B, Supplementary Tables 1 and 2).
 | Figure 5Higher expression level of Pokemon in luminal A type than Basal-like breast cancer tissues, which was just opposite for Smad4. (A, B) The mRNA expression of Pokemon was higher in luminal A type, but lower in basal-like type breast carcinomas, which was just opposite to Smad4. Data was obtained from 2 breast carcinoma database systems.HER2 = human epidermal growth factor receptor 2.
*p < 0.05, †p < 0.01, ‡p < 0.001.
|
DISCUSSION
Pokemon, also known as ZBTB7A, was originally confirmed to be a proto-oncogene due to its function as an inhibitor of tumor suppressor ARF expression [16]. However, recent studies have identified the role of Pokemon as a tumor suppressor in prostate and colon cancer by antagonizing SOX9 transcriptional activity on key target genes such as a tumor invasion involver, MIA, and a long non-coding RNA precursor, H19 [18], as well as by the transcriptional suppression of glycolysis genes [19], suggesting that Pokemon is a context-dependent cancer gene. We previously reported that Pokemon is over-expressed specifically in breast cancer tissue, but not in normal breast tissue, and that Pokemon expression is associated with tumor size, lymph node metastasis, and poor patient survival [14]. In this study, we attempted to identify the role of Pokemon in the growth and proliferation of breast cancer cells and found that Pokemon expression promoted the growth of breast cancer cells. This finding could explain why Pokemon exhibits high levels of expression in breast cancer tissue, since it may act as a key pro-oncoprotein in breast cancer. This result prompted us to further explore the expression pattern of Pokemon in different breast cancer subtypes by using TCGA databases. Pokemon showed the highest mRNA levels in the subtype of Luminal A and the lowest levels in the basal-like subtype. This data indicates distinct expression patterns in different breast cancer subtypes, the function and mechanism of which remains to be elucidated.
In this study, Smad4, as the common mediator of TGFβ signaling, was identified to be one of the downstream genes regulated by Pokemon. Smad4 was found to function as a promoter for TGFβ-induced epithelial-mesenchymal transition in vitro and for bone metastasis of breast cancer cells in vivo [23]. We found that Pokemon suppressed the expression of Smad4 via the repression of Smad4 promoter activity in breast cancer cells. Although no direct binding of Pokemon with the GC-box within the Smad4 promoter sequences was observed, SP1 was found to bind with the GC-box within the Smad4 promoter sequences and enhance the promoter activity, and Pokemon was confirmed to interact with SP1 and abrogate the binding of SP1 with Smad4 GC-box sequence, leading to the suppression of SP1-induced Smad4 promoter activity in MDB-MA-231 cells. This finding might partly illustrate the mechanism by which Pokemon repressed the expression of Smad4, leading to the promotion of the growth and proliferation of breast cancer cells. It was previously reported that Pokemon could inhibit TGFβ-induced transcriptional activities via altering the recruitment of Smad4 co-regulators to TGFβ responsive gene promoters by physically interacting with Smad4 [24]. The physical interaction between Pokemon and Smad4 was revealed to cause the inhibition TGFβ-dependent cell growth arrest in HaCaT cells. Both our and Yang's work provided evidence for the implications of Pokemon in the modulation of TGFβ signaling by interfering the expression and activity of Smad4.
To determine whether Pokemon and Smad4 expression is of clinical relevance, we interrogated databases of gene expression in patient samples from TCGA. Two independent datasets of patient samples indicated that Pokemon mRNA levels were negatively correlated with those of Smad4 in the primary tumors of breast cancer patients. This finding suggests that increased expression of Pokemon might suppress TGFβ-Smad4 signaling to facilitate the growth and proliferation of breast tumors.
In conclusion, in this study, we confirmed that Pokemon plays a key role in promoting the proliferation and growth of breast cancer cells by downregulating the expression of Smad4 through SP1. This highlights the importance of Pokemon as a novel mediator for TGFβ signaling in breast cancer progression.
 XML Download
XML Download