

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a heterogenous disorder characterized by chronic airway inflammation, tissue remodeling and airway hyperreactivity (AHR).1 Based on common clinical presentation as well as on epidemiology, genetics, biomarkers, lung physiology, histopathology and response to therapy, different asthma endotypes have been proposed.1 Allergic asthma is the most frequent asthma endotype and house dust mites (HDMs) allergy is the most frequent among asthmatic patients, which is also one of the greatest risk factors for asthma and AHR.234 Yet, a substantial number of house dust mite-allergic patients (HDM-APs) do not suffer from asthma and have normal bronchial reactivity.56
Many mediators and metabolic pathways are potentially involved in the development of chronic airway inflammation, airway remodeling and AHR in allergic patients.78 Recently, sphingolipids and enzymes involved in sphingolipid metabolism have been proposed to play a role in the pathogenesis of allergic diseases, AHR and asthma.9101112 Sphingolipids play an important role in cell growth, survival, inflammation and tissue remodeling.1314 Sphingosine-1-phosphate (S1P) is of particular interest due to its proinflammatory and anti-apoptotic effects.910 Experimental models of allergic asthma demonstrated that S1P played a crucial role in the development of asthmatic phenotype in different experimental models in mice.1516 Two subcutaneous injections of S1P, 7 days apart resulted in progressively developed airway inflammation and AHR reaching maximum 2 weeks after the last dose.15 Moreover, local application of exogenous S1P aggravated allergen-induced airway inflammation in a mouse asthma model.16 On the contrary, inhibition of sphingosine kinase (SPHK) 1, which is crucial for synthesis of S1P, led to attenuated airway inflammation and AHR in a model of allergic asthma.17 Similarly, application of S1P analogue, which interfered with S1P-S1P receptor interaction, inhibited airway remodeling following repeated allergen exposure in rats.18
Human studies also provided some evidence for a potential involvement of S1P in the pathogenesis of asthma.1920 An increase in concentration of S1P was demonstrated in bronchoalveolar lavage (BAL) fluid 24 hours after segmental allergen challenge.19 In vitro, S1P directly induced pro-remodeling response in human airway smooth muscle cells.20 An S1P plasma-interstitial fluid gradient exists and inflammation-driven increased vessel permeability, as seen in the airways of asthmatic patients, may result in additional influx of S1P to tissue compartment.21
Other sphingolipids also affect processes involved in cell growth, survival and apoptosis.121314 Interestingly, activation of de novo sphingolipid synthesis leads to production of sphinganine (SFA) and ceramide which exert opposite to S1P effects.121314 Therefore, the balance between sphingolipid synthesis and degradation is important for regulation of cell growth, survival, inflammation and tissue remodeling.121314
In the current study, we evaluated the effect of bronchial allergen challenge on plasma concentration of selected sphingolipids in a well characterized group of HDM-APs.
MATERIALS AND METHODS
The study was performed on 33 HDM-APs. All patients reported rhinitis symptoms, while 22 patients reported also asthma symptoms upon exposure to house dust. Sensitization to the HDM components Dermatophagoides pteronyssinuss (Dp) and Dermatophagoides farinae (Df) was confirmed by demonstration of positive skin prick tests and elevated (>0.7 kU/L) serum concentrations of anti-Dp specific immunoglobulin E (IgE). Before the initial visit, none of the patients received allergen immunotherapy or any anti-asthma medication, except sporadic application of short-acting-beta agonists.
The study was approved by the local Ethics Committee (R-I-003/131/2004). All participants provided written informed consent.
Pulmonary function tests
Histamine bronchial challenge was performed as previously described.22 All patients inhaled doubling concentrations of histamine starting from a concentration of 0.125 mg/mL. Forced expiratory maneuvers were performed 90 seconds after fifth inhalation of each histamine concentration. The procedure was continued until either at least a 20% reduction in forced expiratory volume during the first second of expiration (FEV1) or a histamine concentration of 32 mg/mL was reached. Nonspecific bronchial reactivity was expressed as histamine concentration causing 20% fall in FEV1 (PC20).
Bronchial provocation test with aqueous Dp extracts (Allergopharma, Germany) were performed as described before.22 Increasing doses of allergen (0.8, 4, 20, 100, 500 and 2,500 SBU) were administered using a De Vilbis#646 nebulizer attached to a Rosenthal-French dosimeter. Forced expiratory maneuvers were performed 15 minutes after inhalation of each dose of the allergen extract. Allergen inhalations were continued until either at least a 20% reduction in FEV1 (PD20) or a cumulative dose of 5,000 SBU was reached. Subsequently, FEV1 was measured every 15 minutes during the first hour after challenge, every 60 minutes during the next 11 hours and after 24 hours. Specific bronchial reactivity was expressed as the allergen dose causing a PD20. Bronchial challenge with Dp allergen extract was performed on all patients sensitive to HDM allergens.
Exhaled nitric oxide (NO) measurements
Concentration of NO in the exhaled air was measured using a chemiluminescence analyzer NOA 280i (Sievers Instruments, Boulder, CO, USA) according to ATS recommendations as described elsewhere.22 Briefly, each patient exhaled against a fixed expiratory resistance of 16 cm H20 resulting in a constant flow of 50 mL/s. A plateau of NO concentration in the exhaled air at the selected exhalation rate was automatically selected by the computer software. NO measurements were repeated 3 times and the mean value was used for analysis.
Blood samples
Plasma samples were obtained using citrate-theophylline-adenosine-dipyridamole (CTAD) anticoagulation as previously described.22 In addition, EDTA-anticoagulated samples were collected for assessment of complete blood count. The CTAD-anticoagulated blood samples were incubated on ice for 30 minutes and then plasma was separated by centrifugation at 4°C. The supernatants containing platelet poor plasma were aliquoted and stored at −80°C until tested. The samples were collected before bronchial allergen challenge (T0), at 45 minutes (TEAR), 6-8 hours (TLAR) and 24 hours (T24) after administration of the last allergen dose.
Biochemical and immunologic assays
Total IgE and specific IgE were measured in the serum samples using the UniCap system (Pharmacia, Uppsala, Sweden).
Complete blood count, including red blood cell (RBC) and platelet counts, as well as white blood cell (WBC) differential was measured using automated hematological analyzer ADVI-120 (Bayer, Leverkusen, Germany).
Total cholesterol (TC) and high-density lipoprotein cholesterol (HDL-C) were measured using Abbott Diagnostics (Wiesbaden, Germany) on the Architect c8000 analyzer (Abbott Laboratories, Abbott Park, IL, USA).
The levels of ceramide, sphingosine (SFO), SFA, S1P and sphinganine-1-phosphate (SFA1P) were determined as previously described.23 Briefly, lipids were extracted from 250 μL of plasma in the presence of internal standards (10 pmol C17-sphingosine and 30 pmol C17-S1P, Avanti Polar Lipids). An aliquot of the lipid extract was transferred to a fresh tube with preadded 40 pmol N-palmitoyl-Derythro-sphingosine (C17 base; a kind gift of Dr. Z. Szulc, Medical University of South Carolina) as an internal standard, and then subjected to alkaline hydrolysis to deacylate ceramide to SFO. The amounts of S1P and SFA1P were determined indirectly after dephosphorylation to SFO and SFA, respectively, with the use of alkaline phosphatase (bovine intestinal mucosa, Fluka). Free SFO and SFA, dephosphorylated sphingoid bases and SFO released from ceramide were then converted to their o-phthalaldehyde derivatives and analyzed using a HPLC system equipped with a fluorescence detector and C18 reversed-phase column (Varian Inc., OmniSpher 5; 4.6 × 150 mm). The isocratic eluent composition of acetonitrile (Merck, Darmstadt, Germany): water (9:1, v/v) and a flow rate of 1 ml/min were used. All measurements were taken in a blind fashion.
Statistical analysis
Data were analyzed by Student's t test for independent means. Repeated measurements were analyzed using Student's t test for dependent means. For multiple comparisons the Bonferroni correction was applied. Values of P < 0.05 were considered significant. All results are expressed as the means with 95% confidence intervals (CIs). Linear correlation by Pearson was used to estimate correlations between studied parameters. Multiple regression analysis was used to estimation of predictors of individual sphingolipid plasma concentration.
RESULTS
All patients (N = 33) who were allergic to HDM allergens reported rhinitis triggered by exposure to house dust. Twenty-two patients also reported asthma symptoms upon exposure to house dust. The patients developed asthmatic response after bronchial challenge with Dp extract (responders; Rs). The patients who did not complain of asthma symptoms upon house dust exposure (n = 11) also did not develop asthmatic response after bronchial challenge with Dp extract (non-responders; NRs). There was no difference in the demographic characteristics between the 2 groups (Table 1). However, Rs were characterized by greater AHR, peripheral blood eosinophilia and specific IgE concentration. Eleven of the Rs (single responders; SRs) developed isolated early asthmatic response (EAR), while the remaining 11 patients (dual responders; DRs) developed both EAR and LAR after the challenge with Dp extract. The level of sphingolipids examined in our study was in the range previously demonstrated among healthy subjects.24
Table 1
Patient characteristics

The results are presented as means with 95% CI. Geometric means are presented for total IgE, DpIgE, PC20, PD20 and FeNO.
NRs, non-responders; Rs, responders; BMI, body mass index; TC, total cholesterol; HDL-C, high-density lipoprotein cholesterol; IgE, immunoglobulin E; Dp, Dermatophagoides pteronyssinuss; FEV1, forced expiratory volume in 1 second; MEF, maximal expiratory flow; PC20, histamine concentration causing 20% fall in FEV1; PD20, allergen dose causing a 20% reduction in FEV1; FeNO, nitric oxide fraction in exhaled air; RBC, red blood cell; S1P, sphingosine-1-phosphate; SFO, sphingosine; SFA, sphinganine; SFA1P, sphinganine-1-phosphate; CI, confidence interval.
Elevated plasma S1P and SFA1P concentrations are associated with AHR
There was no significant difference between the mean concentration of TC or HDL-C between Rs and NRs (Table 1). In Rs, the mean baseline concentrations of S1P (179 pmol/mL) and SFA (5.9 pmol/mL) were significantly greater than those in NRs (129 and 3.4 pmol/mL, P = 0.0068 and P = 0.0005, respectively) (Table 1). No significant difference in the concentration of ceramide (P = 0.509), SFO (P = 1.0) or SFA1P (P = 0.07) between Rs and NRs was demonstrated (Table 1). Analysis of individual clinical and functional parameters revealed significant associations between plasma concentration of some sphingolipids and lung function parameters. When HDM-APs were divided into 2 groups according to the AHR, significant differences in plasma S1P and SFA1P concentrations were demonstrated. Those with histamine PC20 ≤ 4 mg/mL had significantly greater baseline plasma concentrations of S1P (192 pmol/mL) and SFA1P (27.3 pmol/mL) than those without (126 and 13.4 pmol/mL, P < 0.001 and P < 0.001, respectively) (Fig. 1). In those groups, however, no difference in the mean baseline plasma concentration of ceramide (P = 0.217), SFA (P = 0.118) or SFO (P = 0.979) was demonstrated.
Fig. 1
Plasma concentrations of selected sphingolipids in relation to the severity of AHR in HDM-APs. PC20 ≤ 4; patients with histamine PC20 equal to or less than 4 mg/mL; PC20 > 4; patients with histamine PC20 greater than 4 mg/mL. Only significant differences were indicated.
AHR, airway hyperreactivity; HDM-AP, house dust mite-allergic patient; PC20, histamine concentration causing 20% fall in forced expiratory volume in 1 second.

The baseline plasma concentration of S1P correlated with the baseline lung function expressed as FEV1 (r = −0.563; P = 0.001) or MEF (r = −0.622; P < 0.001) (Table 2). Similarly, a significant correlation was demonstrated between the baseline SFA1P and FEV1 (r = −0.477; P=0.005) or MEF (r = −0.485; P = 0.004). Also, AHR significantly correlated with plasma S1P (r = −0.7451; P < 0.001) and SFA1P (r = −0.5782; P < 0.001) concentration (Table 2). No correlation of the baseline lung function and plasma concentration of any other sphingolipid could be demonstrated.
Table 2
Correlations between individual plasma sphingolipid concentrations and selected functional and biochemical parameters at T0

Bold-faced values are considered significant correlations.
S1P, sphingosine-1-phosphate; SFA1P, sphinganine-1-phosphate; SFA, sphinganine; SFO, sphingosine; FEV1, forced expiratory volume in 1 second; MEF, maximal expiratory flow; FeNO, nitric oxide fraction in exhaled air; PC20, histamine concentration causing 20% fall in FEV1; RBC, red blood cell.
Interestingly, the baseline FeNO correlated neither with S1P (r = 0.307; P = 0.0824) nor with SFA1P (r = 0.242; P = 0.176) plasma concentration (Table 2). However, significant correlations between FeNO and the baseline plasma ceramide (r = 0.505; P = 0.0027) and SFA (r = 0.605; P < 0.001) concentrations were demonstrated. In contrast, peripheral blood eosinophilia correlated with baseline plasma S1P (r = 0.592; P < 0.001) and SFA1P (r = 0.518; P = 0.002), but not with plasma ceramide (r = 0.278; P = 0.117) or SFA (r = 0.338; P = 0.08) concentration (Table 2).
Since ceramide and S1P exert opposite effects on regulation of cell growth, survival, inflammation and tissue remodeling analysis of associations of plasma ceramide/S1P ratio with clinical, functional and immunological parameters were also performed (data not shown). The plasma ceramide/S1P ratio was significantly lower in HDM-APs with histamine PC20 ≤ 4 mg/mL (18.2) than in those without (r = 31.5, P < 0.001). Moreover, the baseline plasma ceramide/S1P ratio significantly correlated with histamine PC20 (r = 0.449, P = 0.019) and baseline lung function expressed as FEV1 (r = 0.351, P = 0.045) or MEF (r = 0.437, P = 0.011) (data not shown).
Peripheral blood eosinophil and platelet counts are independent predictors of S1P plasma concentration
Analysis of the cellular composition of blood samples revealed that the baseline concentration of both S1P and SFA1P correlated with eosinophil (P = 0.0003 and P = 0.002, respectively) and platelet (P = 0.0035 and P = 0.0036, respectively) count, but not with RBC, neutrophil, lymphocyte or monocyte count (Table 2). No significant correlation between the baseline plasma concentration of SFA/SFO or ceramide and any of the cellular blood component was demonstrated (Table 2).
Multiple regression analysis of individual cellular blood components as possible determinants of plasma sphingolipids indicated that peripheral blood eosinophilia (P = 0.003) and platelet count (P = 0.010) were independent predictors of the baseline plasma S1P concentration. Similarly, concentration of SFA1P was determined independently by peripheral blood eosinophilia (P = 0.008) and platelet count (P = 0.009). Concentration of other sphingolipids, including ceramide (P = 0.22), SFO (P = 0.29) and SFA (P = 0.19), could not be predicted based on cellular blood components in the population of HDM-APs (data not shown).
Allergen challenge affects plasma S1P concentration
In DRs, the appearance of EAR was associated with an increase in plasma S1P concentration (from 168 to 215 pmol/ml, P = 0.04), but a decrease in SRs (from 190 to 132 pmol/mL; P = 0.004) (Fig. 2). Moreover, at TEAR an increase in plasma S1P concentration was seen in each DR (100%), while in each SR (100%) a decrease in plasma S1P concentration was demonstrated (not shown). Subsequently, at TLAR, plasma S1P concentration decreased in DRs, while in SRs remained at a decreased level seen at TEAR (Fig. 2). At T24, plasma S1P concentration in SRs but not in DRs was still lower than the baseline value (Fig. 2). No effect of allergen challenge on plasma S1P concentration was demonstrated in NRs (Fig. 2).
Fig. 2
Changes in plasma S1P (A, B and C) and SFA (D, E and F) concentrations during allergen challenge in dual responders (A and D), single responders (B and E) and non-responders (C and F).
S1P, sphingosine-1-phosphate; SFO, sphingosine; SFA, sphinganine; SFA1P, sphinganine-1-phosphate.
*Significantly greater than at baseline; †significantly less than at baseline.

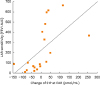
Plasma concentration of SFA, SFA1P, SFO or ceramide did not change significantly during EAR or LAR. However, at T24 plasma concentration of SFA increased in NRs (4.54 pmol/mL, P = 0.04), but did not change significantly in SRs (5.4 pmol/mL; P = 0.55) or DRs (5.7 pmol/ml, P = 0.5) (Fig. 2). Interestingly, the changes in S1P plasma concentration during EAR correlated with the severity of subsequently developed LAR (r = 0.596, P = 0.0071) (Fig. 3).
DISCUSSION
Our study for the first time demonstrated that in HDM-APs, altered intravascular sphingolipid metabolisms, including their phosphorylation and de novo synthesis, are associated with impaired lung function and AHR. In addition, we show that circulating platelets and eosinophils are major determinants of plasma S1P and that SFA1P concentration and allergen-induced asthmatic responses lead to further alterations in intravascular sphingolipid metabolism. An association between asthma and altered sphingolipid metabolism has already been reported.1925 Greater serum baseline concentrations of S1P and SFO were demonstrated in aspirin-exacerbated respiratory disease (AERD) in comparison to aspirin tolerating asthmatics.25 However, the former group did not precisely match the latter in terms of age and severity of AHR.25 The mechanisms regulating intravascular sphingolipid levels are tightly controlled by de novo synthesis and recycling processes.21 The processes effectively operate in both healthy subjects and HDM-APs, so that the baseline plasma concentration of individual sphingolipids in the studied patients remained within the broad range determined previously in a population of healthy subjects.24 Yet, even in healthy subjects, sphingolipid metabolism is affected by age, sex, weight, exercise and blood morphology resulting in significant differences in plasma concentration of individual sphingolipids.21 In contrast to the study by Trinh et al.
25 who found a weak correlation between AHR and serum sphingomyelin concentration, we were able to demonstrate a significant association of AHR with plasma concentrations of S1P and SFA1P. Sphingomyelin represents a reservoir of sphingolipids, but its effects on cell growth and apoptosis are mainly mediated by its metabolite ceramide.121314 It is difficult to speculate if the differences are related to blood processing techniques or truly represent characteristic features of allergic vs aspirin intolerant asthmatic patients. In the blood, platelets constitute a very rich source of S1P which is released upon their activation.21 Therefore, assessment of sphingolipid concentration in plasma, not in serum, seems to better reflect their in vivo metabolism.21 Moreover, it cannot be unequivocally inferred form the study by Trinh et al.
25 whether greater AHR seen in AERD asthmatics is independently linked to altered sphingolipid metabolism as the latter is affected by age and weight of the studied subjects. In our patients, the associations of plasma S1P and SFA1P concentration with lung function and AHR remained significant after controlling for age, sex, body mass index (BMI), TC and HDL-C concentration. Moreover, our study was performed on a homogenous population as all patients were allergic to HDMs. The association between plasma concentration of both phosphorylated sphingolipids with AHR indicates a possible role of enhanced SPHKs activity in the pathogenesis of allergic asthma. In fact, experimental studies demonstrated a crucial role of SPHKs in the development of asthma phenotype in mice.1726 The study with pharmacological inhibition of SPHKs also indicated that the enzymes were crucial for developing AHR, but not airway inflammation in allergic asthma model.26 This is consistent with our findings which indicate that baseline plasma concentration of phosphorylated sphingolipid derivatives, S1P and SFA1P, correlated with AHR, but not with FeNO. On the contrary, elevated plasma concentration of SFA did not correlate with lung function or AHR, but correlated with FeNO. This suggests that lung function impairment and chronic airway inflammation affect different pathways of sphingolipid metabolism. Furthermore, the correlation of plasma SFA with FeNO, but not with AHR, indicates that de novo sphingolipid synthesis may represent a counterregulatory mechanism which is triggered by chronic airway inflammation yet inhibits the development of asthma phenotype in HDM-APs.
Analysis of the dynamic changes in plasma sphingolipid concentration after allergen challenge reinforces the concept of altered sphingolipid metabolisms in the pathogenesis of asthma in HDM-APs. Among all sphingolipids studied, only plasma concentration of S1P increased significantly during EAR in DRs, but not in SRs or NRs. Previously, increased S1P concentrations were demonstrated in BAL fluid 24 hours after segmental allergen challenge in asthmatic, but not non-asthmatic allergic patients.19 Interestingly, the major increase in plasma S1P concentration in the current study was demonstrated much earlier than that reported in BAL fluid.19 An influx of S1P from plasma into bronchial compartment in response to allergen challenge may represent one of the mechanisms responsible for a subsequent increase in its local concentration in the airways. The reported mean concentration of S1P in BAL fluid was more than 20-fold lower than that seen in plasma in our study, which supports a possible S1P influx from plasma to the airway compartment during LAR.19 Interestingly, a systemic administration of S1P was sufficient for developing AHR and asthma phenotype in mice,15 which supports our observations that changes in intravascular sphingolipid metabolism correlated with lung function and AHR. Activated platelets and/or eosinophils, which are major determinants of plasma S1P concentration, may also participate in local increases in S1P concentration in the airways. It was shown that eosinophil number in BAL fluid correlated with S1P level 24 hours after the challenge.19 However, eosinophils were not sufficient to up-regulate S1P synthesis because persistent eosinophilia in BAL 7 days after the challenge was not associated with elevated S1P concentration.19 Platelets are a very rich source of S1P, which can be released from alpha-granules upon platelet activation.21 Allergen challenge leads to intravascular platelet activation in asthmatic patients.22 Moreover, allergen challenge leads to a decrease in the number of circulating platelets and eosinophils possibly due to their sequestration into bronchial tissues.22 It seems therefore that differences in sphingolipid metabolisms between DRs and SRs at TEAR may explain participation of S1P in enhanced AHR associated with the development of LAR in HDM-APs.567
Other experimental studies provide further support for a possible role of S1P in the development of chronic airway inflammation and AHR. In an experimental allergic asthma model, application of FTY720, which down-regulates the activity of S1P receptors S1P1, S1P2 and S1P5, significantly attenuated influx of Th2 cells and eosinophils into the airways of mice sensitized and challenged with ovalbumin.27 In a rat model of repeated allergen exposure, application of FTY720 attenuated airway remodeling.18 S1P could also affect AHR acting directly on smooth muscle cells by inducing a broad range of genes coding for proteins involved in cell proliferation which led to tissue remodeling.2028 Moreover, S1P plays an important role in inflammatory cell chemotaxis, including that of eosinophils, and interacts with its endothelial receptors promoting P-selectin-dependent role of leukocytes.2930 Therefore, it is likely that an increase in plasma S1P at TEAR in DRs participates in influx of inflammatory cells, which in turn is responsible for the development of LAR. This concept is supported by the correlation between changes in plasma S1P concentration at TEAR and the magnitude of the subsequently developed LAR. It may also indicate stronger activation of circulating platelets and eosinophils which are major determinants of plasma S1P concentration. In fact, stronger activation of circulating platelets in response to allergen challenge in DRs in comparison to SRs has already been demonstrated.22 Finally, S1P can directly activate mast cells leading to enhanced bronchoconstriction.3132 We were able to demonstrate the correlation between the severity of allergen-induced bronchoconstriction and changes in plasma S1P, supporting a potential role of S1P in amplification of EAR. Interestingly, we were not able to demonstrate a direct correlation between plasma S1P concentration and FeNO, indicating a possible indirect interaction between chronic airway inflammation and intravascular S1P metabolism. However, the plasma levels of SFA and ceramide correlated with FeNO, which indicates activation of de novo sphingolipid synthesis in response to chronic airway inflammation. Allergen challenge triggered de novo sphingolipid synthesis, but the effect was seen only in NRs at T24; on the contrary, in Rs a tendency for a decrease in plasma SFA was demonstrated. This argues for a protective rather than pro-asthmatic effects of de novo sphingolipid synthesis. Failure to up-regulate de novo sphingolipid synthesis in Rs within 24 hours after the challenge may reflect impaired or delayed compensatory mechanisms in the patients. Orosomucoid like 3 (ORMDL3) which regulates serine palmitoyltransferase activity has recently been implicated in asthma pathogenesis.33 The presence of functional single nucleotide mutations in the ORMDL3 gene which increase its expression is highly correlated with the risk of childhood asthma.33 It seems that elevated ORMDL3 synthesis increases the risk for asthma by depressing de novo sphingolipid biosynthesis.34 If de novo ceramide synthesis in response to allergen challenge were attenuated, the effect of S1P would be enhanced. This concept is supported by detection of a lower ceramide/S1P ratio in the plasma of HDM-APs with greater AHR. It is also consistent with a recently published paper which demonstrates that in mice which overexpress ORMDL3 exogenous S1P dramatically increases AHR and that the effect is much greater than that seen in wild type mice.3536 The small number of patients included in this study did not allow us to perform genetic studies regarding mutations in the ORMDL3 gene. However, the detailed assessment of the patients included as well as multiple longitudinal sampling seems to at least partially offsets the limitations associated with the small number of patients studied. In addition, consistent changes in plasma S1P concentration in DRs and SRs allowed for powerful demonstration of altered sphingolipid metabolism in relation to the development of prolonged airway inflammation associated with late phase asthmatic response.
Increasing body of evidence indicates that asthma is heterogenous and that at least several endotypes based on pathogenetic mechanisms may represent targets for different therapeutic options.3738 Further studies are warranted to demonstrate whether altered sphingolipid metabolisms represent distinct endotypes of allergic asthma or represent characteristic features of allergic asthmatic patients.
In summary, our study provides evidence for altered intravascular sphingolipid metabolisms in HDM-allergic asthmatic patients. Both enhanced synthesis of S1P possibly derived from platelets and/or eosinophils as well as impaired de novo sphingolipid synthesis may participate in the development of asthma phenotype in HDM-APs. Pharmacological modulation of sphingolipid metabolisms seems to be an attractive option for the management of allergic asthma.
 XML Download
XML Download