

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Aspirin-exacerbated respiratory disease (AERD) is characterized by hypersensitivity to nonsteroidal anti-inflammatory drugs (NSAIDs), asthma and chronic rhinosinusitis (CRS) with nasal polyps (NPs).1 Overproduction of cysteinyl leukotrienes (cysLTs) is a hallmark of AERD in the pathogenic mechanisms. CysLTs induce smooth muscle constriction and potentially enhance eosinophil accumulation in the bronchial mucosa.2 Therefore, eosinophilia is commonly found in peripheral blood and upper and lower airway mucosae of AERD patients.3 In addition, AERD is a type 2 immune-mediated airway disease associated with increased expression of Th2 cytokines such as interleukin (IL)-4, IL-5 and IL-13, resulting in persistent eosinophilic inflammation.4 Although many studies have shown evidence that activated effector cells such as eosinophils, neutrophils, mast cells and platelets are involved in the pathogenesis of AERD,5 this review emphasizes recent insights into how eosinophils work in airway mucosa of AERD patients.
SECTION 1: EOSINOPHILS RELEASE MULTIPLE MEDIATORS
A novel molecule released from activated eosinophils may provide a new perspective, as AERD is not fully explained by type 2 cytokines (via Th2/ILC2 responses) or overproduced cysLTs. Extracellular traps from eosinophils composed of DNA and granule proteins are involved in innate immunity and associated with several allergic diseases.6 Moreover, recent studies have revealed that eosinophils from asthmatic patients secrete higher levels of extracellular vesicles, leading to the development and progression of asthma.7 These findings suggest that activated eosinophils contribute to the pathogenesis of AERD through producing several molecules (Table). However, further investigations are needed to understand the role of innate immune responses to activate eosinophils in AERD.
Table
Mechanisms of activated eosinophils in the pathogenesis of aspirin-exacerbated respiratory disease CysLT, cysteinyl leukotriene; IL, interleukin.

![]()
CysLTs
CysLTs, a class of inflammatory lipid mediators, contribute to several characteristic features of AERD. These molecules are derived from effector cells through arachidonic acid metabolism (upon ingesting COX-1 inhibitors such as aspirin and NSAIDs) that oxidizes arachidonic acid to form unstable intermediate leukotriene (LTA4).8 In eosinophils, LTA4 is changed into LTC4 by the enzyme LTC4 synthase and sequentially converted into LTD4.9 Urinary LTE4 (a stable end product) levels, a biomarker for systemic leukotriene production, are significantly higher in AERD patients compared to patients with aspirin-tolerant asthma (ATA) at baseline. Furthermore, these levels even increase 100-fold on aspirin challenge.10 These mediators contribute to eosinophil activation, mucus production, vascular leakage, and edema, which enhance airway inflammation and remodeling in AERD patients.11
Eosinophil extracellular traps
Activated eosinophils release extracellular traps in an NADPH oxidase-dependent manner (associated with reactive oxygen species production), which is distinct from apoptosis and necrosis.12 Many reports have demonstrated that eosinophil extracellular traps are often associated with blood and tissue eosinophilia.1314 Extracellular traps have a function in innate immunity to infectious disease; however, these molecules are cytotoxic enough to induce tissue damage in asthmatic airways.1516 In addition, the percentage of eosinophils forming extracellular traps was significantly elevated under severe airway inflammation.17 Although the pathophysiological function of extracellular traps has not been completely determined, our current study demonstrates that the percentage of eosinophils producing extracellular traps is negatively correlated with baseline forced expiratory volume in 1 second and positively correlated with the levels of eosinophil-derived neurotoxin in serum.18 These suggest that extracellular traps may play a crucial role in severe eosinophilic inflammation and airway obstruction.
Eosinophil extracellular vesicles
Extracellular vesicles are small molecules that contain multiple bioactive proteins, lipids, and nucleic acid, which are important for intercellular communication.19 These membrane-bound carriers are continuously released by several cells and contain different cellular components, depending on their origin.20 Extracellular vesicles have been implicated in the pathogenesis of multiple diseases such as cancer, metabolic disorders, and allergic diseases.21222324 Because of their abundance and unique composition, these molecules have potential as a biomarker for the diagnosis and prognosis of a wide variety of diseases.25 Recently, it has been demonstrated that eosinophils from asthmatic patients secrete a greater quantity of extracellular vesicles than those from healthy control subjects.26 Moreover, production of extracellular vesicles from eosinophils was enhanced in response to inflammatory stimuli such as eotaxin-1 and tumor necrosis factor-α.27 These findings are important to understand the complicated secretory activities of eosinophils underlying immune responses. It is evident that extracellular vesicles participate in allergic diseases with a significant implication in asthma progression. However, further studies are still needed to clarify a specific role of extracellular vesicles in AERD patients.
SECTION 2: EOSINOPHILS INTERACT WITH AIRWAY EPITHELIAL CELLS
Airway epithelial cells represent the first line of the barrier which constantly maintains the mucosal interface epithelium. These cells are involved in innate immunity by expression of diverse pattern recognition receptors against viruses, microorganisms or environmental pollutants.28 Moreover, infiltration of immune cells into the proximity of epithelium results in the induction of adaptive immunity through interactions with epithelial cells.29 Epithelium-derived cytokines such as IL-33, IL-25 and thymic stromal lymphopoietin (TSLP) are implicated in the pathophysiology of asthma.30 In addition to inflammatory features, airway remodeling (changes in the structure and function of epithelium) found in asthmatic patients results in increased resistance of airflow.31 AERD patients present a remodeling process that affects both the upper and lower airways.32 Several molecules released from airway epithelium have been reported to be associated with airway inflammation and remodeling.
Surfactant protein D (SPD)
SPD is a member of the collectin family of proteins mainly produced by airway epithelial cells.33 This molecule is a component of innate immunity within the airways binding to pathogens and inducing phagocytosis by interacting with phagocytic cells.34 In addition, SPD modulates allergic responses through the regulation of eosinophil activation.35 Emerging evidence has demonstrated that SPD is associated with several pulmonary diseases including asthma.36 The SPD level in BALF is significantly lower in patients with severe asthma than in those with mild asthma or healthy control subjects.37 Recently, our group has revealed that the serum SPD level was reduced in AERD patients compared to ATA patients. Moreover, the increased number of eosinophils (LTE4-mediated) in the lungs enhances airway inflammation and remodeling; however, SPD treatment attenuates the symptoms in mice,38 indicating that SPD may have a protective function against eosinophils in the airway inflammation of AERD.
Folliculin (FLCN)
FLCN is an intracellular protein expressed in several inflammatory cells including airway epithelial cells.3940 This molecule has been suggested to be associated with activation of epithelial cells and maintenance of the integrity of the epithelium barrier through regulation of cell-cell adhesion and expression of cell-cell junctions.4142 Impairment of epithelial barrier function (change in junctional complexes) is important in allergic disease including occupational asthma.4344 In AERD patients with NPs, airway epithelium is disrupted by detachment of epithelial cells and damage of intercellular junctions.45 In addition, our recent study demonstrated a significantly higher level of serum FCLN in the AERD group compared to the ATA group.46 Airway eosinophilia found in AERD could increase FLCN release from airway epithelial cells, which enhances epithelial activation and disruption, suggesting that modulation of FLCN may be a potential therapeutic target for AERD.
Periostin
Periostin (a secreted matricellular protein) is produced by airway epithelial cells, leading to eosinophilic inflammation in asthmatic patients.47 It has been demonstrated that periostin is a key player for inducing airway hyperresponsiveness in mice.48 In addition, expression of periostin in airway epithelial cells is up-regulated in the presence of IL-13 (a pleiotropic Th2-cytokine) in vitro.49 IL-13 is released by several immune cells such as eosinophils, mast cells, basophils, and activated T cells.50 Previously, we have shown that the serum periostin level was significantly higher in patients with AERD (severe asthma/eosinophilic asthma) than in those with ATA. In addition, the serum periostin level is positively correlated with blood and sputum eosinophil counts.51 Furthermore, our recent evidence reveals that production of periostin is markedly elevated under the eosinophilic condition and positively associated with the transforming growth factor-β1 level.52 Therefore, periostin may play a crucial role in eosinophilic inflammation and remodeling in the airway of AERD patients.
SECTION 3: THERAPEUTIC APPROACH
For the management of AERD patients, anti-inflammatory medications including inhaled/systemic corticosteroid with or without long-acting beta2-agonist have been prescribed to achieve symptom controls with absolute avoidance from aspirin/NSAIDs.53 Regarding aspects of cysLTs overproduction, several drugs such as leukotriene receptor antagonists and 5-lipooxygenase inhibitors have been applied to suppress the leukotriene production pathways. In addition, a monoclonal antibody targeting the high-affinity receptor binding site on human immunoglobulin E (IgE) was suggested for AERD treatment.54 Furthermore, some ongoing studies on several biologics (targeting type 2 immune responses) have been under clinical trials to control eosinophilic inflammation in AERD patients.
Anti-IL-5 antibody
Previous studies have shown an abundant cytokine and chemokine milieu consistent with type 2 immunity in AERD.4 Some investigations confirmed that there were significant correlations between levels of IL-5 and cysLTs/ECP in NPs and nasal lavage fluid in AERD patients. IL-5 is also elevated in CRS with NPs in AERD patients compared to healthy controls or those with sinusitis without polyps, suggesting that IL-5 plays an important role in eosinophilic inflammation in upper and lower airway mucosae of AERD.55 There are monoclonal anti-IL-5 or anti-IL-5 receptor antibodies such as mepolizumab, reslizumab, and benralizumab, which are suggested effective for severe eosinophilic asthma.56 In the same context, they could also be applicable to control eosinophilic inflammation in upper and lower airway mucosae of AERD.57 Mepolizumab was evaluated in a small, randomized trial of patients with severe NPs, which proposed that it was beneficial for eosinophilic NPs, a key feature of AERD.58 Therefore, inhibition of type 2 response has a potential benefit in managing both conditions, CRS/NPs and AERD.
Antibodies to IL-33 and TSLP
Although eosinophilic inflammation reflects the effect of Th2 cells and allergen-specific IgE, there is a complementary pathway mediated by the innate immune system promoting the similar inflammatory process, especially as an amplifier of Th2 response. Eosinophilic inflammation is initiated by cytokines such as IL-33 and TSLP which are substantially derived from epithelial and other barrier cells damaged by microbes or toxins.5960 IL-33 is an alarmin-like cytokine that activates both myeloid and lymphoid innate effector cells, thereby facilitating production of cytokines like IL-5, IL-13 and IL-9.616263 A recent study demonstrated that IL-33 might be another cytokine playing an important role in the pathogenesis of AERD. The same study revealed that IL-33 was up-regulated in the airway epithelial layer and depended on cysLTs expression in mouse models, suggesting that IL-33 is a component of cysLT-driven innate immune response that promotes activation of mast cells and is attributed to AERD pathogenesis.64 Moreover, IL-33 is a potent stimulus for eosinophil activation that exacerbates airway inflammation.65 TSLP is an IL-7-like cytokine thought to be important in bronchial asthma, atopic dermatitis, and NPs, and induces type 2 cytokine production; therefore, it can activate eosinophils and basophils.6667 A previous study investigated the effect of human monoclonal anti-TSLP IgG2λ in allergic asthmatic patients, suggesting a beneficial effect in reducing bronchoconstriction and airway inflammation against allergen challenge.59 Another report demonstrated that TSLP promoted mast cell-derived PGD2 production and dysregulation of this kind of innate immune system substantially contributes to the pathogenesis of AERD.68 Although these antibodies against IL-33 and TSLP are under clinical trial, they will be future treatment targets for AERD.
CONCLUSION
The clinical feature of AERD is moderate to severe asthma comorbid with CRS and NPs, which is associated with eosinophilic inflammation and cysLT overproduction in upper and lower airways. The pathophysiology of AERD is complicated; however, activation of immune cells (especially eosinophils), dysregulation of the arachidonic acid metabolism and alteration of genetic/epigenetic factors are important mechanisms in AERD patients. In addition, a novel molecule released by eosinophils (extracellular traps and extracellular vesicles) may provide an explanation of more severe airway inflammation and remodeling in AERD. A lower level of SPD (protective function) with higher levels of FLCN and periostin (pathogenic functions) further suggest playing critical roles of eosinophils interacting with airway epithelium, contributing to the pathogenesis of AERD. Therefore, biologics targeting eosinophilic inflammation may have potential benefits to manage AERD patients (Figure).
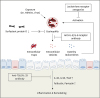 | FigureTherapeutic approach to the management of AERD patients. A leukotriene receptor antagonist attenuates asthma exacerbation caused by cysteinyl leukotriene production. An anti-IL-5/IL-5 receptor antibody inhibits eosinophil activation to release several mediators. An anti-TSLP or IL-33 antibody may potentially reduce type 2 immune responses associated with eosinophilic inflammation.AERD, aspirin-exacerbated respiratory disease; IL, interleukin; NSAID, nonsteroidal anti-inflammatory drug; TSLP, thymic stromal lymphopoietin; SPD, surfactant protein D; CysLT, cysteinyl leukotriene.
|
 XML Download
XML Download