

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Colorectal cancer (CRC) is one of the most common malignancies worldwide, and ranks the second most common cause of cancer-related death for both sexes.1 The morbidity and mortality of CRC have increased in the past few years, especially in developing counties in Asia, such as China.2 Despite advances in the prevention, diagnosis, and treatment of CRC, effective treatment strategies for this disease have yet to be developed.3 Metastasis and relapse are the main causes of failure in the treatment of CRC.4 Therefore, elucidating the molecular mechanisms underlying CRC progression and metastasis is important. Recent evidence supports the involvement of various signaling pathways in the pathogenesis and development of CRC.5 Activation of the Wnt/β-catenin cascade is a common phenomenon in CRC,6 and its downstream target genes can promote tumorigenesis by inducing cell cycle progression and abnormal proliferation.78 The Wnt/β-catenin signaling pathway is involved in the regulation of epithelial-mesenchymal transition (EMT) and cell proliferation, invasion, and migration in CRC. This pathway also plays a vital role in regulating epithelial stem cell self-renewal, and the self-renewal ability of CRC stem cells is important for the initiation of CRC.9 Alterations in the Wnt/β-catenin signaling pathway affect CRC, providing a potential target for treatment. The Janus kinase/signal transducers and activators of transcription (JAK/STAT) signaling pathway, the PI3K/AKT signaling pathway, the MAPK cascade, the p53 pathway, Notch signaling, nuclear factor κB (NF-κB) signaling, and other pathways have also been shown to affect the development of CRC.
Increasing evidence indicates that noncoding RNAs (ncRNAs) play important regulatory roles in various biological processes, including cancer development.10 Long noncoding RNAs (lncRNAs) are ncRNAs longer than 200 nucleotides in length that are not translated into proteins.1112 Recent studies have reported that various lncRNAs act as modulators of carcinogenesis and progression in human CRC.13 LncRNAs are required for CRC cell proliferation and migration,14 are associated with a poor prognosis in CRC,15 and promote colon tumorigenesis.16 LncRNAs exert these effects by regulating various signaling pathways.17 For example, the lncRNA CCAL promotes CRC progression by regulating the Wnt/β-catenin signaling pathway.18 These studies provide new insights with which to improve our understanding of the mechanisms underlying CRC development. Herein, we summarize the latest findings on lncRNAs associated with CRC and highlight associated regulatory mechanisms and signaling pathways that may lead to the development of lncRNA-based approaches in the treatment of CRC.
UNDERLYING MECHANISMS OF LNCRNAS IN THE REGULATION OF CRC
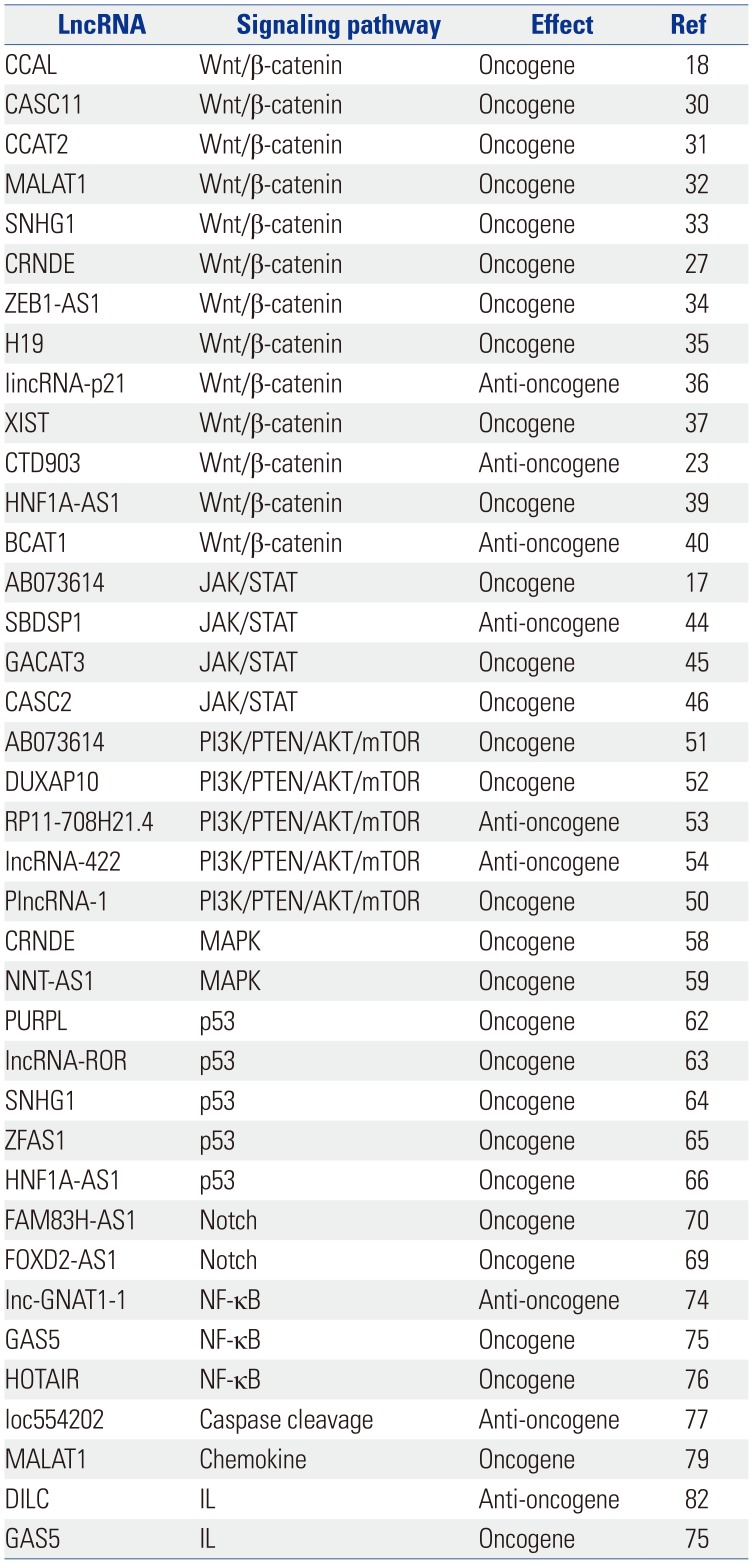
LncRNAs exert effects on gene expression at the transcriptional and post-transcriptional levels via various mechanisms. LncRNAs can bind to DNA, RNA, and proteins to influence transcriptional initiation, RNA stability, or the activity of signaling pathways. LncRNAs serve as a scaffold for the recruitment of transcriptional factors to the promoter region to affect gene expression. Several lncRNAs are involved and play critical roles in the development of cancers, such as malignant proliferation, metastasis, invasion, anti-apoptosis effects, and therapeutic resistance (Table 1). Recent studies have focused on the regulation of lncRNAs in CRC cells through signaling pathways. The underlying mechanisms are addressed in the following paragraphs.
LncRNAs regulate CRC cells through the Wnt/β-catenin cascade
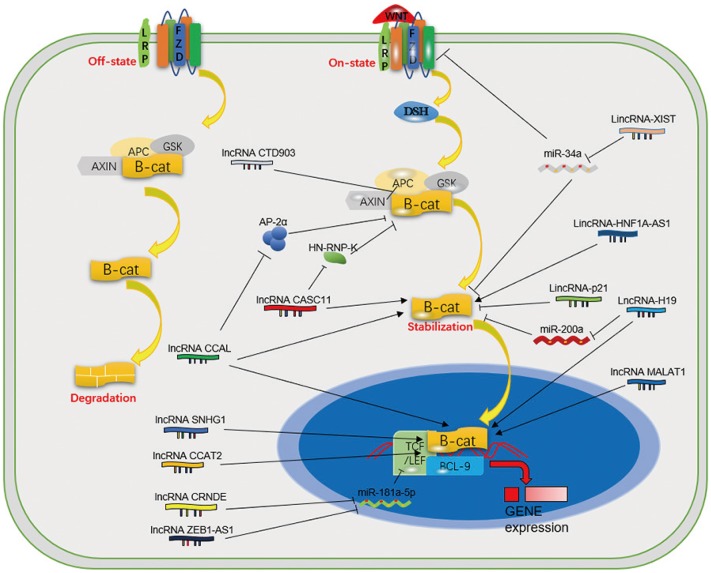
β-catenin affects both carcinogenesis and development. Overexpression of Wnt/β-catenin pathway members is a common feature in CRC.619 Studies suggest that β-catenin plays a role as a transcription factor in concert with TCF1 and LEF1 to activate downstream target genes.20 When Wnt ligands bind to the receptor FZD or LRPs, β-catenin is released from the β-catenin destructive complex, which consist of Axin, APC, and GSK3β. The activation of the Wnt cascade may be induced by mutations of APC.21 Then, β-catenin accumulates and is translocated into the nucleus to activate the downstream genes of the Wnt/β-catenin pathway through hcCF/LEF, and it may continue to induce the process of EMT through increasing the expression of key proteins.2223 Over the past few years, an increasing number of studies have shown that EMT plays a vital role in cancer cell metastasis and invasion.24 Some lncRNAs have proven to affect EMT by or not by Wnt/β-catenin pathway.23242526272829 Several lncRNAs affect the Wnt/β-catenin cascade to regulate CRC (Table 2), and the regulatory mechanisms are listed in Fig. 1.
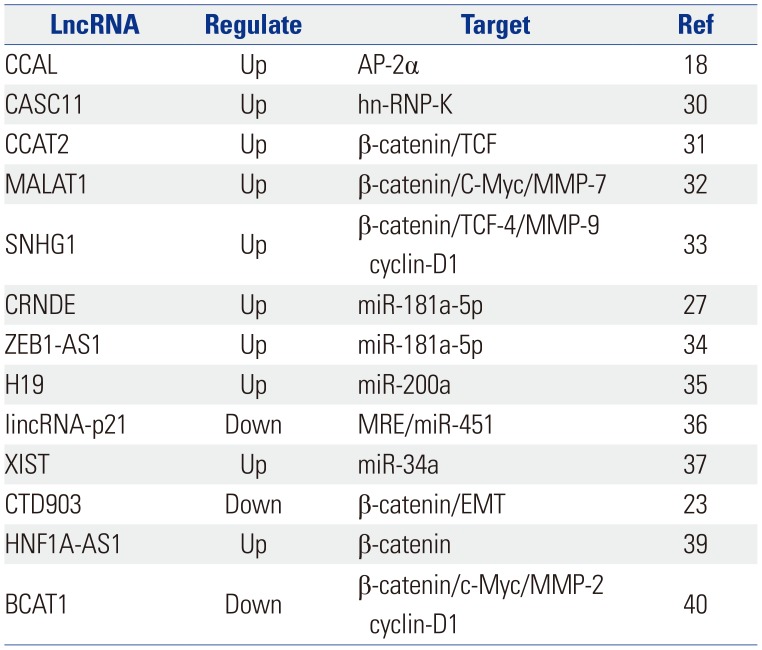
Some lncRNAs, such as CCAL, CASC11, and CCAT2, can bind to proteins or microRNAs (miRNAs) to affect the Wnt/β-catenin pathway indirectly. The lncRNA CCAL can bind to activator protein 2α (AP-2α), which can combine with β-catenin and APC to form a complex, thereby weakening β-catenin/TCF-4 interactions in CRC. The lncRNA CASC11 may activate the Wnt/β-catenin cascade by binding to hnRNP-K, which can form a complex with GSK3β, TCF4, β-catenin, and AXIN to inhibit the degradation of β-catenin. The physical interaction between TCF7L2 and the lncRNA CCAT2 can increase Wnt/β-catenin signaling pathway activity.183031 Some lncRNAs promote the nuclear localization of β-catenin to upregulate the Wnt/β-catenin cascade, such as MALAT1 and SNHG1.3233 MiRNAs are also important targets that help lncRNAs exert effects on signaling pathways by binding to a complex. For example, the lncRNAs CRNDE, ZEB1-AS1, H19, lincRNA-p21, and XIST affect the Wnt/β-catenin signaling pathway by sponging or binding to miRNAs.3435363738 In addition, lncRNAs can change the expression of certain vital proteins to affect the activation of the Wnt/β-catenin cascade. The lncRNA CTD903 may downregulate Wnt/β-catenin signaling and then decrease the expression of TCF, Snail, and Twist to suppress the EMT process, which is closely associated with the development of CRC. The lncRNA HNF1A-AS1 can induce CRC cell growth, invasion, and migration mediated by the Wnt/β-catenin cascade by up-regulating the expression of β-catenin, cyclin D1, and c-Myc. LncRNA-BCAT1 overexpression inhibits β-catenin expression in CRC cells.233940
LncRNAs regulate CRC cells by affecting the JAK/STAT signaling pathway
JAKs and STATs are essential molecules in cytokine signal transduction pathways.41 Despite their long-recognized pivotal roles in immunological responses, recent studies have shown that JAKs and STATs play important roles in cancer initiation and progression.42 The activated JAK/STAT pathway can inhibit cell apoptosis and promote the proliferation and invasion of tumor cells.43 Certain lncRNAs play a role in CRC by modulating the JAK/STAT pathway, acting as competing endogenous RNAs (ceRNAs) or by changing the expression of phosphorylated STAT3. The lncRNAs AB073614 and SBDSP1 contribute to the suppression of STAT3 phosphorylation,1744 whereas the lncRNAs GACAT3 and CASC2 act as ceRNAs for specific miRNAs to indirectly up-regulate SP1 and STAT3 expression.4546
LncRNAs regulate CRC cells by affecting the PI3K/PTEN/AKT/mTOR signaling pathway
The PI3K/PTEN/AKT/mTOR signaling pathway regulates various biological processes in cells, such as gene expression, metabolism, cell growth, and metastasis.47 Although the effects of PTEN are frequently mediated by the negative regulation of the PI3K/PTEN/AKT/mTOR signaling pathway, it can also function as a suppressor independently.48 Recent studies have demonstrated that lncRNAs regulate cancer progression through the PI3K/PTEN/AKT/mTOR cascade, which plays important roles in CRC cell proliferation and EMT.495051 Approximately 40% of malignant tumors are related to alterations of the PI3K/PTEN/AKT/mTOR cascade, which underscores the potential of the PI3K/PTEN/AKT/mTOR pathway as a target for clinical treatment. LncRNAs mainly affect the expression of related proteins to modulate signaling pathways, such as AB073614, DUXAP10, RP11-708H21.4, and lncRNA-422.51525354 In addition, PlncRNA-1 regulates the phosphorylation of PI3K and AKT.50
LncRNAs affect CRC cells through MAPK cascade
Studies have demonstrated that most human cancers are related to a biomolecular event, the activation of Ras/MAPK signaling.55 Activation of the Ras/MAPK pathway promotes cell proliferation, migration, and differentiation.5657 Conversely, inhibition of this pathway in CRC cells suppresses tumor development. The lncRNA CRNDE is important in the regulation of Ras/MAPK pathways. In addition, hnRNPUL2 can increase the stability of CRNDE to act as a vital mediator, thereby activating the Ras/MAPK cascade.58 Wang, et al.59 showed that the activity of the MAPK/Erk signaling pathway is regulated by the lncRNA NNT-AS1 and that activation of the MAPK/Erk signaling pathway can induce EMT in CRC cells, increasing CRC cell proliferation, invasion, and migration.
LncRNAs affect CRC cells through the p53 pathway
P53 is a well-known tumor suppressor that is associated with CRC. Overexpression of p53 promotes cell cycle arrest, apoptosis, and tumor suppression.6061 LncRNAs are regulators and mediators of the p53 cascade, and the effect of many lncRNAs is mediated by regulating the expression of p53. For example, the lncRNAs PURPL, ROR, SNHG1, and ZFAS1 promote tumorigenicity in CRC by suppressing basal p53 levels.62636465 Furthermore, HNF1A-AS1 can repress the miR-34a/SIRT1/p53 feedback loop to down-regulate p53 and promote the metastasis of colon cancer.66
LncRNAs affect CRC cells by modulating the Notch signaling pathway
The Notch signaling pathway is involved in the tumorigenesis of various cancers including CRC.6768 The cascade is observed in the processes of CRC cell proliferation, invasion, and metastasis. In addition, Notch signaling regulates EMT.69 The lncRNA FAM83HAS1 is overexpressed in CRC cell lines and tissues concomitant with the up-regulation of two Notch signaling molecules, Notch1 and Hes1, thereby regulating the progression of CRC.70 FOXD2-AS1 also regulates Notch signaling, acting as a tumor promoter in CRC.69
LncRNAs affect CRC through the NF-κB signaling pathway
The transcription factor NF-κB was identified as a nuclear factor in 1986 and later shown to bind to enhancer elements to exert tumorigenic effects.7172 Under physiological conditions, NF-κB is inhibited by many proteins. When these proteins are phosphorylated, NF-κB is activated and translocated to the nucleus to promote the initiation and progression of CRC.73 Most lncRNAs regulate this cascade by directly or indirectly modulating NF-κB. For example, lnc-GNAT1-1 exerts suppressive functions in CRC through the RKIP-NF-κB-Snail circuit, and lnc-GNAT1-1 knockdown downregulates PKIP and increases NF-κB expression.74 The lncRNA GAS5 can decrease the phosphorylation of NF-κB to activate NF-κB signaling.75 In addition, the NF-κB cascade is associated with 5-fluoropyrimidine resistance caused by lncRNA HOTAIR in CRC.76
LncRNAs affect CRC cells by several other mechanisms
Caspase cleavage cascade
Apoptosis signaling can induce caspase expression, which may affect the progression of CRC. The overexpression of lncRNA loc554202 can promote G1 arrest and cause apoptosis to inhibit CRC cell proliferation, mainly via caspase cleavage cascade.77
Chemokine signaling
Chemokines play an important role in promoting CRC invasion and metastasis.78 The lncRNA MALAT1 promotes the progression of colon cancer through tumor-associated dendritic cells (TADCs), and using neutralizing antibodies or siRNA transfection to block chemokine ligand 5 could diminish the promotion of colon cancer induced by TADCs.79
PERSPECTIVES
LncRNAs account for more than 80% of ncRNAs and generally have a lower transcriptional level than that of chromosome modification, transcriptional regulation, post-transcriptional processing, epigenetic, and nuclear import.11 Recent studies focusing on signaling pathways involved in cellular transformation have identified many cascades as potential treatment targets.47 Several lncRNAs associated with the regulation of signaling pathways have been identified, and lncRNAs have become ideal biomarkers for early diagnosis, therapeutic evaluation, and prognostic prediction in cancer.83 An extensive body of research has demonstrated that a number of cellular signaling pathways initiate and induce the progression of CRC, including the Wnt/β-catenin pathway, the JAK/STAT pathway, the p53 pathway, the MAPK pathway, the PI3K/PTEN/AKT/mTOR pathway, the Notch signaling pathway, and the NF-κB signaling pathway.84 We investigated the effects of lncRNAs on signaling pathways and found that many lncRNAs control CRC properties by targeting different components thereof. Some components of signaling pathways have become attractive therapeutic targets because of their important functions in cancer. For instance, resveratrol can inhibit invasion and metastasis of CRC through MALAT1 because MALAT1 upregulates β-catenin and triggers its transfer to the nucleus.32 However, there are still no candidates for clinical application because of serious side effects, as proper signaling pathway activity is essential to sustaining normal cell survival. How to precisely regulate signaling pathways is challenging. Targeting these regulatory lncRNAs may be an alternative approach with which to achieve precise modulation of signaling pathways.
CONCLUSION
We summarized multiple functions and signaling pathways related to lncRNAs in CRC cells and highlighted their potential clinical applications as novel diagnostic and prognostic biomarkers and therapeutic targets. Our review provides new insights with which to better understand CRC and points toward the potential for lncRNA-based therapeutic strategies for CRC.
 XML Download
XML Download