

 PDF
PDF ePub
ePub Citation
Citation Print
Print



THE PROBLEM
Ischemic heart disease can lead to myocardial infarction (MI) and is one of the major health concerns worldwide. Myocardial ischemia and/or ischemia-reperfusion injury (IRI) can damage heart muscle, reducing its ability to pump efficiently. A sudden and/or severe blockage of a coronary artery can lead to a MI. Myocardial ischemia can also cause serious arrhythmia and sudden death. Cardiac remodeling that involves inflammation, infarct expansion and subsequent scar formation follows the ischemic injury. The remodeled (dilated) heart requires neuroendocrine activation to maintain systemic hemodynamics, but chronic neuroendocrine activation exacerbates structural remodeling and functional abnormalities.1)2) After MI, inflammatory cells enter the heart to clear dead tissue and then promote scar formation.3) Damaged myocytes in the MI border zone, that have uncoupled from the undamaged myocardium, usually die, and then the infarct expands, the heart dilates and a persistent increase in wall stress is imposed on the surviving myocardium. Patients with a large scar burden with dilated hearts can develop heart failure ultimately leading to premature death.
OVERVIEW FOR CELL-BASED THERAPIES
Cell-based therapies for cardiac repair and regeneration have emerged recently as a promising alternative to existing pharmacological and surgical interventions. Currently, cardiac treatment is primarily designed to be ‘damage-limiting’ modality, which is unable to prevent adverse remodeling and scar formation. In contrast, adoptive transfer of reparative stem cells into an injured myocardium can improve cardiac pump function although the exact mechanisms remain debatable. There are many published studies that have tested a variety of stem cell types to see if they have some capacity for cardiac repair after MI.4)5)6) A variety of adult stem cell types that might repair the injured heart have been tested in animal models. These studies have shown that transplantation of autologous cardiac-7)8)9)10) or bone marrow-derived11)12) stem cells induced pluripotent stem cells and direct reprogramming of endogenous non-stem cells into cardiogenic phenotypes13)14) have some capacity to improve cardiac function after injury. Some of these preclinical successes have been translated into early stage clinical trials.15)16)17) Early stage clinical trials have largely focused on autologous (derived from the patient) stem cells due to their ease of isolation and lack of immunogenicity. These trials suggest that both bone marrow-18)19)20) and cardiac-derived15)16)21) cells offer modest functional benefits when transplanted after cardiac injury. The outcomes of these trials have been somewhat variable, but the overall effects of autologous stem cell therapies are a small improvement in cardiac structure and function. Importantly, due to the time requirements to prepare autologous cell therapeutics, the therapy is delivered after endogenous repair has begun and often after mature scar has formed.
The fundamental mechanisms of stem cell mediated repair are still largely unknown and highly controversial.16)22)23) Some early studies in animal models suggested that differentiation10)11)12) of injected cells into new cardiac myocytes is a major mechanism of cardiac repair. Studies with c-Kit+ cardiac and bone marrow derived stem cells suggested that these cells could robustly (trans) differentiate into new cardiac myocytes when injected into the infarcted heart.11)24) Since replacing cardiac myocytes lost from ischemic insult is the ultimate goal of effective cell therapy, these results were extremely promising. However, the majority of studies from multiple independent laboratories using a variety of methods have not confirmed these early results. Most of reliable recent studies suggest that differentiation of these stem cells into cardiac myocytes is a rare occurrence at best and is not a major mechanism of stem cell mediated improvements in cardiac structure and function.25)26) The consensus of recent studies is that paracrine factors from injected stem cells enhance cardiac repair in the infarct border zone through a variety of mechanisms. The stem cells induce neovascularization, activate endogenous stem cells, and modify the post MI inflammatory response, which promote resolution of inflammation and reduce scar formation. They also enhance a small amount of new myocyte generation that appears to be resulted from preexisting myocytes proliferation in the host. Collectively these processes induce the beneficial effects of cell therapy.8)9)23)27)
Why there is a need for new/potent stem cell source?
Both the heart and bone marrow are thought to contain stem cell niches, in a dedifferentiated state, that contain cells with the capability to differentiate into the cardiac or mesenchymal lineage.11)28)29)30) In these environments, the pluripotent state of stem cell is thought to be supported by complex signaling interactions with surrounding cells, such as those found in the stromal lining of the bone marrow cavity. It is now clear that stem cells from the bone marrow have a very limited capacity to form cardiac myocytes31) and c-Kit+ cardiac stem cells (CSCs) are difficult to isolate and expand, and do not appear to have significant cardiac myocyte lineage capacity.32) The ability of these cell types to repair the heart by differentiation into new cardiac myocytes has not been validated. Therefore, we explored the development of a novel stem cell type “cortical bone derived stem cells (CBSCs)”. CBSCs are more easily isolated and have a greater capacity to repair damaged heart tissue, primarily by improving wound healing after ischemic injury.33) Previous studies from our group and others suggest that the cortical bone, rather than the bone marrow, might be a source of stem cells with more “primitive” characteristics.33)34) We have shown that CBSCs can be easily obtained through routine bone biopsy procedures, are negative for most markers of the hematopoietic lineage, and express a unique profile of cell surface markers that are distinct from typical mesenchymal stem cells (MSCs).35) We have recently documented that delivery of “autologous” CBSCs in mouse MI models can improve cardiac structure and function with a small amount of differentiation of the CBSCs into new cardiovascular cells. The small cardiogenic potential of CBSCs was not to be primarily responsible for their beneficial therapeutic effect. These experiments suggested that reparative effect of CBSCs involve secreted paracrine factors that improve wound healing after ischemic injury.
WHAT ARE CORTICAL BONE DERIVED STEM CELLS?
A number of CSC and extra CSC types have been used for cell-based therapies for cardiac repair. As discussed above, various limitations such as lack of cardiomyogenic differentiation, susceptibility to ischemia, and decreased paracrine signaling, warrant identification of new cell type for cardiac cell therapies. We have recently identified CBSCs, a novel stem cell population from the bone stroma.35) CBSCs possess a unique cell surface marker profile when compared to 2 cell types used in different phases of clinical trial i.e., MSCs and cardiac derived stem cells (CDCs). CBSCs highly express CD55, integrin β3 (CD61), CD82, NT5E (CD73), integrin β4, and endoglin, compared to CDCs and MSCs.33) Concurrently, CBSCs express extremely low levels of CD59 compared to CDCs and MSCs, further highlighting their uniqueness. Expression of CD96 was extremely low in CBSCs and CDCs, compared to MSCs. Similarly, CD248 was expressed on CBSCs and CDCs. All 3 cells types expressed CD276, CD109, and were negative for PTPRC (CD45) and CD11b. Simultaneously, CBSCs are clonogeneic marking their stemness nature.33)
CORTICAL BONE DERIVED STEM CELLs FOR MYOCARDIAL REPAIR
The effects of CBSCs delivery showed improvement in structure and function of hearts post-MI (Figure 1).
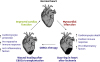 | Figure 1CBSCs for wound healing after MI.CBSCs = cortical bone stem cells; MI, myocardial infarction.
|
Small animal models studies
Extensive studies in small animal models using C57BL/6 mice explored the potential beneficial effects of injecting CBSCs into the border zone of animals after inducing a MI with permanent occlusion of the left anterior descending artery (LAD). Our results showed that CBSCs have beneficial effects on the structure and function of the heart 6 weeks after cardiac injury.35) Animals with MI that received a CBSCs transplantation had improved survival and cardiac function, and attenuated adverse left ventricular remodeling compared with both saline-injected MI controls and animals that received CDCs. Cardiac function assessment using echocardiographic myocardial strain analysis showed that CBSCs-treated animals not only had improved global function, but also had greater myocardial contractile function at the MI border zone relative to CDC-injected hearts, suggesting that the stem cells improved contractile function in the region where they were injected.35) These studies showed that “autologous” CBSCs can transdifferentiate into a few new vessels and small myocytes. However, the improvement of structure and function was not directly related to the amount of neovasculogenesis or myocytes, suggesting that paracrine effects of these cells are the major mechanisms of wound healing in the injured heart.
Large pre-clinical animal studies
After thorough characterization of CBSCs in a small rodent model where we confirmed that CBSCs reduce ventricular remodeling and improve cardiac function after MI, the next logical step was to test it in a large pre-clinical animal model that could be applied in real post-MI patients. This approach required the use of “off-the-shelf” allogeneic porcine CBSCs. This study explored the short- and long-term benefits of delivering CBCSs into the border zone of mini-swine with IRI.36) A NOGA® mapping system (Biosense Webster, Diamond Bar, CA, USA) was used to assess electric conductance within the heart and determine the interface between viable and nonviable tissue, without opening the chest. The system was also used to guide catheter-based transendocardial injections. The 30 injection sites were observed in explanted hearts with gross visualization of fluorescent microspheres within the myocardium in all animal for the short-term study (72-hour) and a long-term study (3-month) cohorts.36) The animals were infused with EdU for the first 3 days after MI to identify cells with newly formed DNA. A large number of EdU+ cells were observed 3 days after CBSCs transplantation. The majority of these EdU+ cells were not myocytes, but predominately made up of von Willebrand factor+ cells, CD45+ cells, and smooth muscle actin+ cells, hinting involvement of other wound healing process rather than just regeneration of myocytes. CBSCs treatment in animals showed 2-fold increase in the percentage of EdU+ nuclei in the border zone of the injected area indicating modulation of wound healing response after CBSCs injection. The long-term study showed that CBSCs delivered after IRI in mini swine reduced left ventricular dilation and preserved cardiac functional reserve.36) These CBSCs treatments also reduced scar size, likely by reducing the number of dying myocytes and via immunomodulation. CBSCs injection also inhibited hypertrophic remodeling.
WHAT IS CARDIAC WOUND HEALING?
There are 2 kinds of ‘wounds’ that have clinically-high relevance to the cardiovascular system. One is the rupture of an atherosclerotic plaque and the other is MI. The first dominant healing process after MI occurs in early inflammatory phase. In this phase, an influx of immune cells that release pro-inflammatory mediators or paracrine factors occurs and dead tissues are cleaned out.37) The second reparative phase leads to increased matrix synthesis, proliferation of fibroblasts and scar formation. The dying cardiomyocytes at the border zone of MI secrete a variety of pro-inflammatory proteins called chemokines, which are primarily designed to elicit responses from bone marrow derived cells and play a critical role in inflammatory response to cardiac injury. Additionally, studies have documented an activation of cytokine signaling in the infarcted myocardium. Many different pro-inflammatory cytokines that have functional pleiotropy and redundancy are increased in the injured myocardium and affect biological processes and exert similar or opposite effects on the same cell type. The analysis of these processes may be of major therapeutic importance, especially for reparative therapeutics involving stem cells. For the development of therapeutics to improve wound healing, it will be imperative to understand if and how stem cells modulate these different phases of wound healing. Here, we will focus on how CBSCs modulate the early/late inflammatory phase and help in preventing the expansion of infarct size. It is important to appreciate how CBSCs interact with macrophages (MФs), T cells and other immune cell types, which will improve understanding of wound healing process after CBSCs infusion in IRI models.
There is a growing consensus in this field that MФs play an important role in cardiac regeneration.38) Biology of MФs has undergone a paradigm shift recently with the discovery of resident tissue MФs.39) Three distinct cardiac MФs populations exist based on differential cell surface marker expression. Two of the MФs subsets originate from embryonic progenitors and are maintained through self-renewal rather than monocyte differentiation. The third cardiac MФs subset is derived from circulating blood monocytes and mainly enters the heart during a pro-inflammatory response following acute and chronic cardiac injury.40) Recent evidence suggests that embryonic derived MФs possess increased ability to phagocytose dying/dead cardiomyocytes after ischemic injury in parallel with secretion of anti-inflammatory cytokines such as interleukin 10 (IL-10). In contrast, high expression of pro-inflammatory genes and a low phagocytic activity of monocyte derived MФs appears to contribute to cardiac inflammation. Strategies that allow expansion of MФs derived from embryonic progenitors while simultaneously suppressing pro-inflammatory MФs may be effective in reducing cardiac inflammation and could enhance cardiac repair. Concurrently, the role of T-cells in cardiac ischemic injury is thought to be very important but has been largely overlooked. However, recent data indicates that CD4+ T-cells influence cardiac repair response in the heart following ischemic injury.41) Interestingly, activated CD4+ T-regulatory cells enhance repair after MI by transforming MФs into a pro-reparative state. These recent studies highlight the importance of understanding immune cell response after injury and their potential interaction with transplanted cells to move the cell therapy field forward.
DO CORTICAL BONE DERIVED STEM CELLs HAVE A ROLE IN WOUND HEALING?
An emerging idea that stem cells can modulate the immune response in the injured heart has generated enthusiasm in the field. Cardiac damage results in release of pro-inflammatory cytokines that triggers a cascade of events. The immune response promotes the healing of damaged necrotic tissue generated by the ischemic insult. Injured myocardium has shown to release pro-inflammatory chemokines, which further stimulate the recruitment of immune cells to the injured tissue. Neutrophils and MΦs act as the first responders to cardiac injury.42) The next phase involves attraction of T lymphocytes to the site of injury that secretes pro-inflammatory factors, which increase influx of other types of immune cells including T, B and antigen-presenting cells (APCs) to the site of injury. However, prolonged intense inflammation can result in further damage, eventually leading to organ failure. Immune modulatory properties of stem cells may promote resolution of inflammation and facilitate tissue repair. Previously, mesenchymal stem cells (MSCs) have been well reported to play positive beneficial role in immune modulation and are known to be immuno-privileged.43) Therefore, we aimed to study if CBSCs possess any markers that might be involved in mediating/altering the immune response to injury. CBSCs expressed extremely low levels of IL-1, secreted phosphoprotein 1, and IL-18 compared to MSCs and CDCs. These factors are known to play a pro-inflammatory role and trigger T, B cells and APCs response. IL-1 is mainly produced by activated MФs and neutrophils and is known to play central role in mediating immune responses. Low expression of these factors suggests CBSCs can play a positive role in modulating the immune response, which might lead to improve wound healing cardiac repair after infarction. Concurrently, CBSCs also increase tumor growth factor β (TGFβ) expression, which can inhibit T lymphocyte proliferation. It has been demonstrated that anti TGFβ antibodies can restore T lymphocyte proliferation. Collectively, these data suggest that CBSCs might have a potential role in regulating immune suppression when delivered into the damaged heart. Our preliminary findings demonstrate that CBSCs derived paracrine factors enhance the number of M2 MФs in CBSCs injected animals. Simultaneously, the finding of increased expression of pro-inflammatory factors rather than anti-inflammatory factors has further strengthened the evidence that CBSCs can modulate immune response (unpublished data). Ischemic injury to the adult myocardium is characterized by myocyte death, activation of inflammatory signaling and scar formation. Cell-based therapies are being tested as an approach to enhance cardiac repair but the results to date have been modest. Therefore, exploring other properties that can modulate multiple biological processes is necessary to move the field forward.
CAN PARACRINE SIGNALING FROM STEM CELLS AUGMENT HEALING PROCESSES?
Well-characterized stem cells that are shown to enhance cardiac repair could lead to more effective therapies for patients with ischemic heart disease. In our previous work35) we have determined that CBSCs secrete various factors that enhance cardiac repair after ischemic insults. These factors include angiopoietin-1 (Ang-1), basic fibroblast growth factor (bFGF), hepatocyte growth factor (HGF), insulin-like growth factor-1 (IGF-1), platelet-derived growth factor (PDGF), stem cell factor (SCF), stromal-derived factor-1 (SDF-1), and vascular-endothelial growth factor (VEGF). HGF and IGF-1 are thought to be cardioprotective: HGF has cytoprotective, anti-apoptotic and pro-angiogenic effects,44)45)46) while IGF-1 can inhibit apoptosis and may stimulate growth and proliferation of stem cells.9)22)24)45)46) SCF, and SDF-1 are thought to stimulate stem cell function: SCF, the ligand for the c-kit receptor,47) may stimulate stem cell homing,48) while SDF-1 is a chemotactic ligand that induces stem cell proliferation and homing to the site of injury.22)46)49) Ang-1, bFGF, PDGF and VEGF all promote angiogenesis: Ang-1 induces vascular cell migration and enhances stability of newly-formed vasculature,46)48) bFGF induces proliferation of endothelial and smooth muscle cells22)45)46)48) and fibroblast, PDGF stimulates smooth muscle cell proliferation22)46)48) and VEGF induces endothelial cell proliferation and tube formation.22)45)46)48)49) Our most recent work in mice35) suggests that stem cell mediated secretion of VEGF is primarily involved in increased blood vessel density after MI. Collectively, our published work support the idea that CBSCs have the characteristics to secrete a host of paracrine factors which are regulating the wound healing process after myocardial ischemic injury. These paracrine factors can target multiple biological processes including modulation of immune response, fibrosis and new vessel formation, which leads to smaller infarct sizes in animals treated with CBSCs. As we suggest above, the consensus among many studies8)50) published to date is that the beneficial effects of stem cells in the damaged heart primarily come from paracrine/chemokine factors of injected cells that modify healing.51) Bone marrow-derived stem cells appear to act primarily through paracrine stimulation of new blood vessels and/or through differentiation into vascular cells rather than transforming to myocytes.52) Therefore, it is necessary to revisit the basic biology behind stem cells in order to develop better understanding on how they work and how their effects can be improved after transplantation.
CORTICAL BONE STEM CELLS RESTRICT INFARCT SIZE AFTER MYOCARDIAL ISCHEMIA
One of the most striking findings in our small and large pre-clinical animal studies is that CBSCs treated animals showed a significant smaller infarct sizes after cardiac injury. Cardiac fibroblast plays a vital role in wound healing after myocardial injury and affects multiple aspects of wound healing response including deposition of extracellular matrix proteins to wound angiogenesis and scar maturation. Following acute MI, cardiac fibroblasts in the heart become activated and rapidly proliferate. In rodent hearts, peak cardiac fibroblast numbers are achieved within 7–14 days after permanent ligation of the LAD53) and within 3 days of IRI54). Many laboratories using multiple stem cell types have reported the persistence or presence of stem cells during the same time window after transplantation. Consequently, it is extremely important to study the role and interaction of transplanted cells with the fibroblast to uncover how scar formation and maturation occur after cell therapy. As mentioned above CBSCs treated animals showed a significant smaller infarct sizes, there is a need to better understand their relationship with the cell type (fibroblast) that is a key player in scar formation and maturation as an important arm in wound healing.
FUTURE DIRECTIONS
Adoptive transfer of stem cells into post MI heart exposes them to pro-inflammatory cytokines, chemokines and immune cells accumulated as result of injury to the myocardium. The dying cardiac tissue induces a constellation of pro-inflammatory cytokines and chemokines that trigger an inflammatory response and subsequent recruitment and activation of immune cells to the site of injury. A number of studies have confirmed that regardless of the cell type used, a large population of the donated stem cells are lost very early after transplantation. Whether this early loss of transplanted stem cells is largely dependent on the hostile post MI environment remains undefined. Ideally, stem cells that can withstand the onslaught of detrimental factors and at the same time possess the ability to manipulate immune/inflammatory responses to transform the cardiac environment into a pro-reparative state would, in our view, be a major advance in the field. CBSCs demonstrate increased properties of proliferation and survival. Work to date documents that transplantation of CBSCs into injured myocardium improves cardiac structure and function. CBSCs appear to have some ability to form distinct cardiac cell lineages but largely appear to release cardioprotective paracrine factors at the site of injury for better wound healing after injury.
In this concluding portion we discuss some of the major questions that remain unanswered and are necessary before we can apply cell therapy to masses. First, given the diversity of biological processes involved in cardiac wound healing it is important to understand interaction of stem cells with immune cells, fibroblasts, and endothelial cells to understand the basic biology for wound healing and how stem cells could influence these processes. How and what stem cells target both in the early and in the late phase of wound healing? In particular how CBSCs target and modulate wound healing.
Secondly, to validate the presence of this novel potent stem cell type in human bone stroma. Subsequently, a through characterization of human CBSCs with the follow up large animal studies is imperative to move application of CBSCs therapy forward. We would expand the study of CBSCs in post IRI/MI hearts to a large (pig) animal model using human CBSCs. This next step is important for a number of reasons. First, the physiological properties of hearts from rodents and large mammals are fundamentally different, making translation from rodents to humans problematic.55) One example is that the manner in which rodents and large mammalian myocyte regulate diastolic and systolic (Ca2+) is distinctly different.56)57) There are many other examples of these differences.58) Therefore, therapeutics shown to have beneficial actions in rodents may or may not have these same effects in larger animals and with the cells isolated from human bone stroma.
Other issues that need attention are route of delivery and doses of the therapeutics. We have to develop and test therapeutics that mimics the situation that is usually present in people suffering from an MI and then having their coronary vessels opened via catheterization or thrombolytics. Currently, we employ a mapping system (NOGA®; Biosense Webster-guided injections into specific regions of the heart or injection via the coronary vasculature) that could be readily translated into humans. However, we still have to optimize the dose and time of CBSCs delivery that most efficiently improves wound healing. Thus, further research is necessary to better understand the interaction of inflammation and cardiac healing after stem cell delivery. This will allow us to choose better targets at better time points for clinical intervention.
 XML Download
XML Download