

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a complex disorder that involves both inflammation and skin barrier dysfunction. Reduced skin barrier function, specifically caused by abnormalities in filaggrin expression and lipid synthesis, can lead to the release of thymic stromal lymphopoietin (TSLP) from keratinocytes, inducing T helper 2 (Th2) immune response1. Subsequently, Th2 cytokines, such as interleukin (IL)-4 and IL-13, and pro-inflammatory cytokines, such as IL-8 and tumor necrosis factor-alpha (TNF-α), are secreted, resulting in a dermal inflammatory reaction.
Both IL-4 and IL-13 cause abnormalities in lipid production and epidermal differentiation23 as well as inhibition of antimicrobial peptide synthesis in the epidermis4, which allows microbial invasion that is damaging the epidermal barrier. Furthermore, IL-8 (CXCL8), a pro-inflammatory cytokine that directly affects immune cells, including polymorphonuclear cells, is secreted by immune cells after antigen sensitization56. Keratinocytes are considered another major source of IL-8 that is overexpressed in AD skin67, leading to a vicious cycle of repetitive inflammation and deterioration of the barrier function.
Adipocytes and sebocytes secrete adiponectin, an adipokine, which reaches a concentration of 3~30 µg/ml in the plasma89. Adiponectin functions as signal molecule via at least two adiponectin receptors, AdipoR1 and AdipoR2, on target cells10. Because it acts primarily as a metabolic mediator of insulin sensitivity and glucose homeostasis, adiponectin has been implicated in various diseases, especially metabolic disorders such as diabetes and obesity and cardiovascular diseases101112. It has been demonstrated that the adiponectin level in the plasma is negatively associated with the accumulation of body fat, particularly visceral fat, and that the plasma adiponectin levels are low in obese individuals13. In addition, adiponectin plays a role in controlling inflammation during immune responses and, therefore, it has emerged as an important therapeutic target for chronic inflammatory disorders due to its pleiotropic activities in regulating inflammation and immune response, and vascular homeostasis141516. Several studies have reported on the association between psoriasis and adiponectin in relation to inflammation and metabolic abnormalities as well as the possibility to exploit the adiponectin signaling mechanism as a therapeutic target171819.
Recent advances in three-dimensional (3D) skin equivalent models have facilitated the development of AD-like 3D skin equivalent models by subjecting the cells to a treatment with major cytokines involved in AD pathogenesis62021. These models can be used to study the inflammation and barrier function of the epidermis. Specifically, epidermal equivalent models of AD consisting of keratinocytes enable studies that are focused on epidermal physiological phenomena, such as morphogenesis, proliferation, and differentiation and the properties of stratum corneum lipids.
In the present study, we investigated the effects of adiponectin, which has anti-inflammatory and cytoprotective effects, on inflammation, lipid synthesis, and epidermal differentiation in the 3D epidermis equivalent model exposed to the inflammatory condition of AD.
MATERIALS AND METHODS
Reagents and antibodies
Recombinant human full-length adiponectin protein was obtained from Biobud (Seongnam, Korea). We obtained antibodies against AdipoR1 and AdipoR2 from Santa Cruz Biotechnology (Santa Cruz, CA, USA); filaggrin from Abcam (Cambridge, UK). Polyinosinic-polycytidylic acid [Poly(I:C)] was purchased from Sigma-Aldrich (St. Louis, MO, USA), and TNF-α, IL-4, and IL-13 were from R&D Systems (Minneapolis, MN, USA).
Immunohistochemical staining
Subjects were not recruited specifically for this study to obtain human skin samples, but the usage of skin samples from subjects who had previously provided informed consent for research, was approved by the Dankook University Hospital Institutional Review Board (DKUH-2015-02-017-001) and the analysis was performed retrospectively.
For examination by light microscopy, full-thickness human skin specimens or epidermal equivalents were fixed in 10% formalin solution. After dehydration with a graded series of ethanol and xylene, the tissues were embedded in paraffin and cut into 4-µm thick sections. After de-paraffinization and rehydration, hematoxylin and eosin (H&E) stains were used for histological examination.
Standard immunohistochemical (IHC) protocol was followed after de-paraffinization and rehydration. Subsequently, each 4-µm section in 10 mM sodium citrate (pH 6.0) was heated in a microwave oven for antigen retrieval, after which endogenous peroxidase activity was blocked with blocking serum solution (Dako, Carpinteria, CA, USA). Thereafter, adjacent sections from the same paraffin block were incubated with the primary antibodies: AdipoR1, AdipoR2, and Filaggrin (diluted 1:100) at 4℃ overnight, followed by incubation with secondary antibody. Signals were developed with two-component high-sensitivity diaminobenzidine (DAB) chromogenic substrate (Dako Cytomation Liquid DAB Substrate Chromogen System; Dako). The slides were covered with cover glass and then examined using a light microscope (BX43, Olympus, Tokyo, Japan)
The 3D in vitro AD-like human epidermal equivalent model
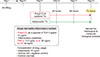
The 3D in vitro epidermal equivalent model based on cultured HEEs was exposed to an AD-inducing inflammatory cytokine cocktail to create atopic conditions according to a previously published method (Fig. 1)62021. The 3D human epidermal equivalents (HEEs) were supplied by TEGO Science Inc. (Seoul, Korea) on days 9~10 of the experiment. Briefly, to construct epidermal equivalents, keratinocytes were seeded into culture inserts. After culturing the cells for 3 days, the constructs were cultured at the air-liquid interface for 10 days. On day 11 after the air-lift culture of constructs, the inflammatory cocktail (10 µg/ml Poly(I:C), 3.5 ng/ml TNF-α, 30 ng/ml each IL-4 and IL-13) was supplemented (AD-like model, AD-HEEs) or not supplemented (normal-HEEs) to the culture medium until harvest after 72 hours on day 14. The constructs were incubated at 37℃ and 5% CO2, and the medium was changed every 2 or 4 days. The culture medium was supplied by TEGO Science Inc.
Quantitative real-time reverse transcriptase polymerase chain reaction analysis
The total RNA from HEK was extracted using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA), and the amount of RNA was determined using the Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Carlsbad, CA, USA). The total RNA (1 µg) was reverse transcribed using M-MLV reverse transcriptase (Promega, Madison, WI, USA) according to the instruction of the manufacturer. The quantitative real-time reverse transcriptase polymerase chain reaction (qRT-PCR) was performed with SYBR Green PCR master mix (Applied Biosystems, Foster City, CA, USA) using Quantstudio 3 (PCR Instrument system, Thermo Scientific Inc., Waltham, MA, USA). The PCR gene expression level was normalized to GAPDH expression. Quantification was performed by the critical threshold (CT) method. The primer sequences for the qRT-PCR are presented in Table 1.
Statistical analyses
The data are expressed as mean±standard error. The data were analyzed by the paired Student's t-test and analysis of variance (ANOVA) with Bonferroni's post hoc correction for comparison between the mean of two numeric values. Statistical significance was defined at p-value <0.05 and the statistical analyses were performed using PASW Statistics ver. 18.0 (IBM Co., Armonk, NY, USA).
RESULTS
Expression of adiponectin receptors decreased in the lesions of AD
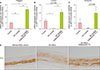
To investigate the changes in the expression of AdipoR in the lesional skin of AD, IHC staining was performed with anti-AdipoR1 and AdipoR2 antibodies in three skin samples each from young normal adults and same-aged adults with AD. The expression of AdipoR1 and AdipoR2 was reduced in the lesional epidermis of patients with AD, suggesting that the adiponectin signal transduction activity may be weakened in the AD epidermis (Fig. 2).
Adiponectin treatment reduced AD-specific inflammatory mediators in the inflammatory HEEs with AD features
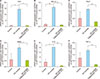
By the qRT-PCR analysis, the changes in the expression of major inflammatory mediators in AD were evaluated after adiponectin treatment in the 3D AD-like model. Carbonic anhydrase II (CAII) and neuron-specific NEL-like protein 2 (NELL2) have been reported as epidermal molecular markers of AD owing to their specific up-regulation in the atopic lesional epidermis2122. Therefore, the increase in the expression of CAII and NELL2 in our AD HEEs indicated that it is similar to the epidermis of AD. This increase was subsequently decreased to the control level in the reconstructed epidermis co-treated with adiponectin for 72 hours (Fig. 3A, B). In keratinocytes, the increased expression of two cytokines, TSLP and IL-8, typically secreted during Th2 immune responses, was also significantly normalized after adiponectin treatment (Fig. 3C, D). Moreover, the expression of TNF-α, a pro-inflammatory cytokine, and human beta defensin 2, an anti-microbial peptide, was similar to that of TLSP (Fig 3E, F). These results suggested that adiponectin has the effect of counteracting the inflammatory responses of AD.
Adiponectin reduced the histological inflammatory response
In the H&E-stained sample used to evaluate the degree of morphological changes, the AD-HEE group (treated with AD cocktail only) showed intercellular spongiosis, and lack of keratohyalin granules and stratum corneum formation (Fig. 4B). In contrast, the adiponectin-treated AD-HEEs demonstrated less spongiosis and relatively well-developed corneal layer with basket-weave structure (Fig. 4C), and they stabilized to a relatively normal epidermis (normal-HEEs) (Fig. 4A).
Adiponectin increased the expression of lipogenic enzymes and transcription factors in AD-like inflammatory conditions
We further evaluated the epidermal lipid synthetic potential closely related to skin barrier function. The expression of the rate-limiting enzymes for fatty acid, cholesterol, and ceramide biosynthesis (fatty acid synthase [FAS], HMG CoA reductase [HMGCR], and serine palmitoyl transferase [SPT], respectively) was decreased in AD-HEEs but significantly upregulated by adiponectin treatment (Fig. 5A~C). In addition, the mRNA expression of the transcription factors encoding the genes related to the biosynthesis of fatty acid or cholesterol, sterol regulatory element binding protein-1c (SREBP1c), and -2 (SREBP2), was restored to normal control level in adiponectin-co treated AD-HEEs (Fig. 5D~F). These results suggested that the lipid synthesis ability of reconstructed epidermis, which was reduced in the AD condition, can be restored by the adiponectin effect.
Adiponectin potentiated the epidermal differentiation markers in the inflammatory state of AD
The mRNA expression of filaggrin, involucrin, and loricrin, the major epidermal differentiation proteins, was significantly decreased due to AD inflammation, but the expression was increased above the normal control level after adiponectin treatment (Fig. 6A~C). Further, the expression of filaggrin protein in HEEs, which is important for the pathogenesis of AD, was further examined by immunohistochemistry. Adiponectin-treated AD-HEEs demonstrated the increased expression of filaggrin in the viable layer of epidermal equivalent compared with that in AD-HEEs (Fig. 6D). Thus, adiponectin might control epidermal differentiation abnormalities under AD inflammatory conditions.
DISCUSSION
The present study was conducted to investigate the effects of adiponectin on Th2 inflammatory cytokine-induced 3D HEEs. The results revealed that adiponectin inhibits the expression of major inflammatory mediators from keratinocytes, enhances lipid synthesis ability, and promotes epidermal differentiation, which might help to improve AD.
CAII is involved in the maintenance of cellular pH, water transport, and ion homeostasis; therefore, it could also contribute to pathological changes that are characteristic for AD, such as spongiosis23. NELL2 is a neural tissue-specific epidermal growth factor-like repeat domain-containing protein22. The pathophysiological role of NELL2 in AD has not been elucidated. However, it is known that the expression of NELL2 is elevated by Th2 cytokines (IL-4 and IL-13) similar to CAII expression and is specifically increased in AD lesions2224. Thus, CAII and NELL2 are considered molecular epidermal markers of AD25.
The 3D model used in this study was established according to the existing model described in previous studies, which showed that the increase of CAII and NELL2 mRNA is a relatively specific event for AD epidermal inflammation6212223. Similar to the observations documented in these previous reports, we also observed an increase of inflammatory cytokines related to AD, such as TSLP and IL-8, and the increase of CAII and NELL2, as well as AD-associated histopathological changes in our 3D model. Therefore, we reconstructed the AD-like inflammatory conditions in our in vitro experiments.
Adiponectin exerts anti-inflammatory actions on several cell types including macrophages, endothelial cells, cardiomyocytes, and fibroblasts9. Adiponectin reduces lipopolysaccharide-induced expression of TNF-α in cultured macrophages through the inhibition of nuclear factor-κB (NF-κB) signaling and also exhibits anti-inflammatory effects via the direct modulation of the macrophage phenotype262728. The expression of TNF-α and matrix metalloproteinase-12 in macrophages in the lung decreased by adiponectin treatment and the infiltration of CD4+ T lymphocytes into atherosclerotic lesions was attenuated via the suppression of T cell chemoattractants in macrophages2930. In endothelial cells, adiponectin also attenuates the inflammatory response in vascular walls by reducing adhesion molecules and IL-6 expression31. Furthermore, adiponectin attenuates TNF-α expression in cardiomyocytes and fibroblasts, leading to reduced cellular apoptosis32. Therefore, adiponectin has a protective effect on various organs, including the heart, vasculature, and lung, by balancing the excessive inflammatory response, such as the inhibition of TNF-α induced NF-κB activation.
In the case of psoriasis, a typical chronic inflammatory skin disease, serum adiponectin levels were found to be inversely correlated with disease severity in patients with psoriasis3334. This finding is convincing while considering a series of reports that suggest psoriasis is associated with metabolic syndromes as well as reports of low blood adiponectin levels in obese individuals and patients with diabetes19. Moreover, it has been recently reported that adiponectin level in the skin tissue and subcutaneous fat decreased in patients with psoriasis, and adiponectin regulates psoriasis-induced skin inflammation by suppressing IL-17 production from human CD4+ or CD8+ T cells17. These observations suggested that adiponectin signaling is a candidate mechanism to explain the relationship between psoriasis and metabolic disorders, and it plays a role in controlling cutaneous inflammation.
In relation to allergic inflammatory disorders, including AD, interest in adipokines is increasing in an effort to identify the mechanism of increased prevalence or severity of metabolic problems, such as obesity, as in psoriasis. The serum and tissue transcriptional levels of adiponectin were lower in patients with AD (especially in patients with extrinsic AD) than in normal subjects, and they were inversely correlated with the total IgE level353637. Furthermore, in asthma, an airway allergic condition, lower levels of adiponectin were associated with uncontrolled or greater risk for asthma3839. However, there is also a report that implicates the negative effects, that is, adiponectin does not protect against the development of inflammation in asthma and might exacerbate the disease via its anti-TH1 inflammatory effects, allowing increased TH2 differentiation and more severe allergic response40. Although there is only limited data available to exactly determine the effect of adiponectin, considering the present results that the expression levels of inflammatory mediators, such as TSLP, IL-8, and TNF-α, were reduced in AD-HEEs by adiponectin treatment, it can be assumed that the normalization of the serum adiponectin level by lowering its level in obese patients by weight control might improve skin inflammation and epidermal barrier function.
We also found decreased expression of adiponectin receptors in the epidermis of patients with AD, which might result from the suppression exerted by the inflammatory cytokines. If this is true, further studies are necessary to confirm whether the reduction in skin inflammation severity by anti-inflammatory agents leads to the normalization of receptor expression.
Adiponectin increased the mRNA expression of differentiation proteins (filaggrin, loricrin, and involucrin) and the major epidermal lipid biosynthetic enzymes (FAS, HMGCR, and SPT) in the AD-like epidermis. The differentiation promotion, especially the increase in filaggrin expression is consistent with the findings of previous studies that adiponectin directly upregulated filaggrin expression in normal human epidermal keratinocytes through the sirtuin 1-mediated pathway4142.
Accordingly, for an effective AD therapy, it is expected that the activation of the adiponectin pathway might enhance epidermal differentiation and barrier function as well as attenuate the inflammatory response of keratinocytes. Therefore, the development of an adiponectin-like agonist of adiponectin signals or receptors might be beneficial for the treatment of AD. Further studies are needed to validate our findings and elucidate the underlying mechanisms.




 XML Download
XML Download