

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Hair aging is a complicated process involving various factors. The study of hair aging is divided into two main streams1. One is an intrinsic biological process at cellular level that is directly linked to hair growth. For hair growth and cycling, interaction between human dermal papilla cells (hDPCs) and mesenchymal stem cells in hair follicles and various growth factors are essential2. The other one is an extrinsic process that happens outside the scalp1. Extrinsic process is highly related to the cuticle layer. The cuticle exists at the outermost part of the hair. It protects the hair shaft from external mechanical stress. The cuticle layer is damaged by external stress such as ultraviolet rays, smoking, and malnutrition. It is known that when the cuticle layer is damaged, aging process of the hair can be initiated easily3.
In terms of structure of hair cuticle which protects hair from extrinsic stimuli, ceramide is the most important intracellular lipid that constitutes the lipid layer of hair cuticle along with fatty acid and cholesterol4. Ceramide is a component of intercellular lipids that are present in hair cuticles to protect and strengthen internal components of hair. Moreover, ceramide is a crucial second signal molecule that regulates various signaling pathways, including cell cycle, cellular senescence, and apoptosis5.
Previous studies have shown that ceramide has an antagonistic effects for cell growth and cell survival6. However, recently it has been reported that ceramide can affect hair loss prevention and hair growth enhancement7. In particular, ceramide synthase 4 (CerS4) plays an important role in the maintenance of epidermal stem cell homeostasis and regulation of the hair cycle by regulating Wnt signal and BMP signal8. Although ceramide can affect Wnt/BMP balance in vivo8, the effect of ceramide on growth pathway such as Wnt/BMP balance in vitro has not been confirmed.
Therefore, the objective of this study was to investigate the effect of ceramide on hDPCs at cellular level. To investigate the effect of existing natural C8-ceramide (N-octanoyl sphingosine) and ceramide-like synthetic compound on hair growth, we evaluated the hair growth pathway including Wnt/β-catenin and BMP signaling changes after stimulation with ceramide. Three types of ceramide were used in this experiment: C8-ceramide and two newly synthetic ceramides (oleyl and stearyl ceramide). These newly synthesized ceramides are different from C8-ceramide in that they have higher solubility through double bonds and carbon chain length.
MATERIALS AND METHODS
Ceramide material preparation
C8-ceramide was purchased from Cayman Chemical (Ann Arbor, MI, USA). Oleyl ceramide was synthesized by reacting one equivalent of 2-dodecenyl succinic anhydride and oleyl amine in methylene chloride at room temperature (RT) for 30 minutes. The reaction mixture was then extracted with ethanol and water and recrystallized to a pale yellow paste, with a purity of 95% and a yield of 98%. Structure of the compound was confirmed by 1H-NMR and LC-MSMS spectrometer. 1H-NMR spectra were recorded on a ZEOL 400 MHz FT-NMR spectrometer in CDCl3. LC-MSMS spectra were recorded on a Shiseido HTS HPLC system and an AB SCIEX API 3200 triple quadruple mass spectrometer. LC-MSMS separations module with electrospray ionization was carried out in positive ion mode. Stearyl ceramide was synthesized in the same manner using stearyl amine (Fig. 1A). All ceramides were dissolved in ethanol to obtain concentration of 1 mM and store at −20℃.
Cell culture
hDPCs were purchased and cultured in Follicle Dermal papilla cell growth medium (PromoCell GmbH, Heidelberg, Germany) supplemented with 10% fetal bovine serum and 1% penicillin and placed in an incubator at 37℃ with 5% CO2.
Cell viability assay (MTT assay)
The viability of hDPCs was evaluated by 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay. Briefly, hDPCs were seeded into 24-well plates at density of 1.0×104 cell per well. After 48h of incubation for hDPCs to reach confluency, three kinds of ceramides (C8-ceramide, oleyl, and stearyl ceramides) at various concentrations of 10 µM, 100 µM, 500 µM, 1,000 µM and 5,000 µM were used for treatment for 24 hours, 48 hours, and 72 hours under 1% penicillin conditions. After incubation, 100 µl of MTT solution was added to each well and the hDPCs were then incubated at 37℃ for 4 hours. After the medium was aspirated, 400 µl DMSO was added to each well. The absorbance of the plate was measured at wavelength of 540 nm using an ELISA plate reader.
Real time polymerase chain reaction
RNA was isolated from cultured hDPCs using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA) and cDNA synthesis was performed using a cDNA synthesis kit (Takara, Shiga, Japan) according to the manufacturer's instructions. The cDNA was used for real-time polymerase chain reaction (PCR) which was carried out with SYBR Green (Bio-Rad, Hercules, CA, USA). Sequences of primers used in real-time PCR are listed in Table 1.
Western blot analysis
The protocol for western blot analysis was described in a previous report9. Briefly, protein lysates from hDPCs were prepared using radio immunoprecipitation assay cell lysis buffer containing protease inhibitor cocktail. The cell lysate was collected by scraping after removed cell debris from lysed cells. Protein concentration was determined using a Bradford assay. Proteins were separated by 10% SDS-polyacrylamide gel electrophoresis and blotted onto polyvinylidene fluoride membrane. The membrane was blocked with 5% BSA/TBST reagent for 1 hour at RT. It was then incubated with primary antibodies against total ERK, total Akt, phosphorylated-ERK, Akt, and GSK3β, Cyclin-D1, β-catenin (Cell Signaling Technology, Beverly, MA, USA), Bcl-2, BAX, and β-actin (Santa Cruz Biotechnology Inc., Dallas, TX, USA) overnight at 4℃ on a rotary shaker. The blotted membrane was then visualized with an AI600 Bio-Image analysis system (GE Healthcare Life Science, Chicago, IL, USA).
Growth factor array
Synthetic ceramides were screened for their effects on the expression of growth factors by using Human Growth Factor Antibody Array G1 (RayBiotech Inc., Norcross, GA, USA) in accordance with the manufacturer's protocol.
Immunofluorescence staining
Cultured hDPCs were seeded on 4-well chamber slides at density of 5×103 cells per well (SPL Life Science, Pocheon, Korea). After starvation, hDPCs were treated with C8-ceramide, oleyl, and stearyl ceramides at concentration of 1,000 µM for 24 hours. After aspirating the medium, cells were rinsed with 1X PBS three times for 5 minutes each time. Then hDPCs were fixed with 4% paraformaldehyde for 15 minutes and permeabilized with 0.1% Triton X-100 for 10 minutes. After washing with 1X PBS, cells were blocked with 1% BSA in 1X PBS for 1 hour at RT on a shaker. They were incubated with anti-rabbit β-catenin (Cell Signaling Technology) primary antibody diluted at 1:100 in blocking buffer and incubated at 4℃ overnight. On the next day, they were incubated with Alexa Fluor 488 labeled goat anti-rabbit secondary antibody (Invitrogen) diluted at 1:200 in PBST at RT for 1 hour in the dark. These stained cells were mounted with mounting medium containing DAPI to counterstain nuclei. Cells were then observed with a fluorescence microscope (Axiovert 200; Zeiss, Oberkochen, Germany).
RESULTS
Viability of hDPCs after treatment with ceramides
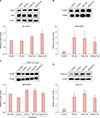
To investigate the effect of synthetic ceramides (Fig. 1B) on viability of hDPCs, we performed MTT assay. Firstly, optical concentrations of oleyl and stearyl ceramides and C8-ceramide as a positive control were determined. hDPCs were treated with various concentrations (0 µM, 10 µM, 100 µM, 500 µM, 1,000 µM, and 5,000 µM) of oleyl and stearyl ceramides, and C8-ceramide for 72 hours. Results showed that oleyl and stearyl ceramides inhibited cell viability at 5,000 µM (Fig. 1C). However, viability of hDPCs was increased after treatment with C8-ceramide (110%), oleyl ceramide (109%), and stearyl ceramide (102%) at 1,000 µM. Based on these results, 1,000 µM was chosen as the proper concentration of ceramides for further experiments.
Ceramides regulates Bcl-2/BAX and ERK/Cyclin-D1 in hDPCs
We examined expression levels of Bcl-2 and BAX known as apoptosis regulator genes by western blot analysis. Results showed that Bcl-2 expression was significantly enhanced by treatment with ceramides compare to that in the control group. However, treatment with ceramides showed no significant effect on BAX expression (Fig. 2A). When the ratio of Bcl-2 to BAX was compared, there was no significant difference between C8-ceramide treatment group and the control group. However, in groups treated with the two newly synthesized ceramides, the ratio of Bcl-2 to BAX was increased 1.22-fold in the group treated with oleyl ceramide and 1.53-fold in the group treated with stearyl ceramide. Such increases were statistically significant.
ERK is also an important factor involved in cell survival and proliferation1011. We examined the effect of ceramides on phosphorylated-ERK in order to determine if our synthetic ceramides could affect mitogen-activated protein kinase (MAPK) signaling involved in the growth of hDPCs. Protein expression level of phosphorylated-ERK was increased following treatment with ceramides (Fig. 2B). PD98059, an ERK inhibitor, is known to regulate the proliferation of hDPCs. hDPCs were pretreated with ERK inhibitor PD98059 (10 µM) for 1 hour and then incubated with ceramides for 24 hours.
Treatment of PD98059 reduced the overall activation of ERK compared to the untreated group. In Fig. 2C, phosphorylated-ERK expression was abolished in the PD98059 treated group compared to the untreated group. Also, we tested whether the ceramides affected phosphorylation tendency to aggravate. As a results, in the ceramide-treated group, the expression of phosphorylated-ERK was suppressed compared with Fig. 2B, but the expression was increased as compared with the control group.
Our results revealed that the expression level of Cyclin-D1 was increased after treatment with C8-ceramide, oleyl, and stearyl ceramide (Fig. 2D). These results indicate that our synthetic ceramides could regulate proliferation of hDPCs by regulating Bcl-2/BAX ratio and ERK/Cyclin-D1.
Ceramide treatment upregulates growth factors including epidermal growth factor family
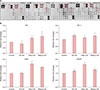
Growth factors were assessed using growth factor membrane microarray (RayBiotech Inc.) according to the manufacturer's instructions. As shown in Fig. 3A, treatment with oleyl and stearyl ceramides increased growth factors levels overall. Epidermal growth factor (EGF) family members such as amphiregulin (AR), EGF, EGF receptor (EGFR), and insulin-like growth factor 1 (IGF-1) are known to stimulate nuclear translocation of β-catenin.
We confirmed that the IGF-1, EGF and EGFR expressions were increased in response to ceramides-treatment (Fig. 3B). AR expression was decreased by C8-cer treatment, but oleyl and stearyl ceramide-treatment increased AR expression.
Ceramides activates β-catenin pathway in hDPCs
We focused on the β-catenin pathway involved in hair growth. The level of phosphorylated-Akt was significantly increased by treatment with ceramides compared to that in the control group (Fig. 4A). Our results also showed that oleyl and stearyl ceramides had a reversal effect on LY294002, an Akt inhibitor. They induced a decrease in phosphorylation of Akt (Fig. 4A). Our results showed that protein expression level of phosphorylated-GSK3β was increased by treatment with ceramides including C8-ceramide, oleyl, and stearyl ceramides (Fig. 4B).
Western blot results showed that β-catenin expression was increased in groups treated with ceramides compared to that in the control group (Fig. 4B). Additionally, our immunofluorescent staining results confirmed the expression of β-catenin in the nucleus. Interestingly, the expression of β-catenin in the nucleus was very weak in the control group untreated with ceramides whereas the expression of β-catenin in the nucleus was significantly increased in groups treated with synthetic ceramides. In particular, its expression in the nuclei and cytoplasm in groups treated with synthetic ceramides (oleyl and stearyl ceramides) was significantly increased compared to that in the control group (Fig. 4C). These results suggest that our synthetic ceramides could induce β-catenin expression.
Wnt/β-catenin and BMP signaling in hDPCs
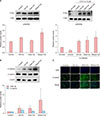
Wnt signaling is known as the master regulator of hair growth and hair cycle while β-catenin is a key transcription factor activated by Wnt signaling12. BMP signaling is known to regulate stem cell activation during hair regeneration13. To determine whether our synthetic ceramides might affect Wnt/β-catenin and BMP signaling, we conducted real-time PCR analysis. As shown in Fig. 5, treatment with stearyl ceramide increased Wnt signaling such as mRNA expression level of wnt3a (2.1-fold), wnt5a (1.8-fold), and wnt10b (6.1-fold) (Fig. 5A). Treatment with stearyl ceramide also significantly increased β-catenin signaling, including mRNA expression level of β-catenin (5.5-fold), AP-1 (3.2-fold), Lef-1 (3.8-fold), and Axin2 (2.2-fold) compared to the control group (Fig. 5B). Moreover, treatment with oleyl ceramide increased mRNA expression level of wnt3a, wnt5a, and wnt7a (Fig. 5A), as well as β-catenin, AP-1, Lef-1, and Axin compared to the control group (Fig. 5B).
The mRNA expression of BMP2 and BMP4, which are known as BMP target gene, were downregulated by oleyl ceramide treatment. Stearyl ceramide treatment also induced a decrease in BMP4 expression (Fig. 5C). Our data support that our synthetic ceramides could regulate Wnt/β-catenin pathway by inhibiting BMP target genes.
DISCUSSION
Ceramide is central component of cellular membrane. It is a key player in many signaling pathways involved in cell cycle arrest and differentiation14. Previous studies about effects of ceramide on hair have focused on developing cosmetics to protect hair shaft15. However, recent studies have suggested that ceramide promotes hair lengthening, leading to increased anagen follicles in mice and human organ culture system7. These results indicate that ceramide might not only play a role as hair shaft protector, but also act as hair growth stimulator. Consistent with previous studies, our results showed that the two newly synthesized ceramides could stimulate hair growth at cellular levels.
In the present study, we demonstrated that synthetic ceramides could increase proliferation of hDPCs accompanied by increased Bcl-2/BAX and phosphorylated-ERK/Cyclin-D1 expression levels. We also observed that expression levels of β-catenin stimulated genes such as phosphorylated-Akt/GSK3β and AR/EGF/EGFR/IGF-1 were increased after treatment with ceramides. In addition, oleyl and stearyl ceramides could upregulate the expression of Wnt/β-catenin signaling and downregulate BMP2 and BMP4 (BMP2/4) expression. Taken together, our results suggest that synthetic ceramides have hair growth stimulating potentials.
Studies about hair growth have shown that the proliferation of hDPCs is accompanied by increased duration of anagen phase16. Hair follicle size is determined by the volume of hDPCs while hDPCs volume depends on the number of cells17. Hence, we evaluated the effect of synthetic ceramides on hDPCs proliferation. hDPCs were treated with various concentrations of ceramides. Our results confirmed that synthetic ceramides at 1,000 µM could increase viability of hDPCs. These results indicate that these synthetic ceramides might contribute to the proliferation of hDPCs.
Apoptosis is mainly processed through the intrinsic death pathway18. The most important regulators of apoptosis process are members of the Bcl-2 gene family. Bcl-2 protein is known to possess anti-apoptotic ability to obstruct a variety of apoptotic signals. BAX protein is another member of the Bcl-2 family. It plays a pro-apoptotic role by stimulating apoptosis function, unlike Bcl-2 protein19. It has been reported that cellular Bcl-2/BAX ratio represents regulation of cell survival20. Also, Bcl-2/BAX ratio was considered important in the hair cycle. It is known that the expression of Bcl-2 is dependent on the cycling of hair follicle, and apoptosis is the central program of the hair cycle21. Our results revealed that BAX expression did not change while Bcl-2 expression was significantly increased in groups treated with synthetic ceramides. Bcl-2/BAX ratio was increased in oleyl or stearyl ceramide treated group compared to that in the control group.
Ceramide activation may either activate or inhibit the MAPK pathway depending on the cell type22. MAPK signal is known to regulate various cellular responses such as cellular proliferation, cell differentiation, development, and apoptosis23. ERK is known as one of MAPK signaling pathways involved in cell survival and growth10. Also, ERK pathway was known stimulate hDPCs proliferation24. Cyclin-D1 is also a key protein that promotes cell cycle (G1-S phase) progression. An increase in cyclin-D1 expression is required for cell proliferation to occur25.
In this study, we examined the effect of synthetic ceramides on phosphorylation of ERK and Cycin-D1 expression. Results showed that treatment with oleyl and stearyl ceramides increased phosphorylation of ERK and induced expression of Cyclin-D1 protein. Previous studies have shown that high concentration of ceramide in cells induces apoptosis6. However, our results showed Bcl-2/BAX ratio and ERK/Cyclin-D1 levels were increased by treatment with our synthetic ceramides. Although the underlying apoptotic effect of ceramide on hDPCs is not fully understood yet, our results show that synthetic ceramides may stimulate proliferation of hDPCs by regulating Bcl-2/BAX ratio and ERK/Cyclin-D1 pathway.
Hair growth involves various transcription factors. EGF family members are known to stimulate nuclear translocation of β-catenin and upregulate the expression of Wnt10b and β-catenin levels. Especially, IGF-1 is known as an important effect on hair growth and extended the anagen phase generally26. The Wnt/β-catenin signaling pathway is known as a master regulator for hair growth and hair follicle morphogenesis12. We investigated Akt/β-catenin signaling pathway to elucidate the molecular mechanism involved in the effect of ceramides relate to β-catenin activation. To activate β-catenin translocation into nucleus, activation of Akt is needed. Akt is involved in signal transduction pathways such as inflammatory mediator, glycogen metabolism, and proliferation apoptosis27. Activation of Akt results in phosphorylation of several substrates such as GSK-3β. GSK-3β is an active enzyme negatively regulated by Wnt pathway and Akt-induced phosphorylation. In the Wnt pathway, GSK-3β phosphorylation and inactivation will lead to the accumulation and nuclear translocation of β-catenin28. Activated β-catenin induces Wnt signaling activation. Therefore, β-catenin is an essential molecule in Wnt signaling. As shown in Fig. 4, ceramides-treatment increased phosphorylated GSK3β via activation of Akt. Finally, β-catenin expression in the nucleus was increased by ceramide-treatment. Our findings are consistent with results of Kwack et al.16 reporting that minoxidil can increase phosphorylation levels of Akt/GSK3β and β-catenin expression.
The balance between Wnt/β-catenin pathway and BMP pathway is important for hair growth cycling29. Axin2 and Lef-1 are directed target genes of the β-catenin pathway. Axin2 plays a role in regulating β-catenin level. Cytoplasmic protein is activated when Axin2 binds to β-catenin. Lef-1 is known as the binding site for β-catenin to accelerate directional differentiation of hair follicle stem cells30. Wnt3a and Wnt7a are inductive signals to maintain hDPCs in an anagen state31. Wnt5a is a key target gene of Shh in hair follicle morphogenesis32. Wnt10b, also known as hair follicle's growth stimulator, promotes hair follicle regrowth by upregulating Wnt/β-catenin signaling pathway related factors including Axin2 and Lef-133. Previous studies have also reported that Wnt10b/β-catenin signaling causes regeneration of hair follicles and proliferation of hair matrix cells3334. As shown in Fig. 5, treatment with synthetic ceramides upregulated expression of β-catenin target genes such as Lef-1, AP-1, and Axin2. Especially, Wnt10b and β-catenin expression levels were significantly increased by treatment with stearyl ceramide. These findings indicate that treatment with synthetic ceramides has potential to affect hair growth.
BMP signaling can promote telogen. It has been reported that inhibition of BMP signaling induces Wnt signaling and initiates hair growth early13. BMP2/4 activation can elaborately regulate the development of hair follicle bulge stem cell activation and growth during hair cycle35. As shown in Fig. 5, treatment with oleyl ceramide downregulated expression levels of BMP target genes (BMP2/4). These results suggest that downregulating BMP2/4 expression could help hair growth via early activation of Wnt signaling.
In conclusion, our study demonstrated that synthetic ceramides had positive effects on hair cell proliferation by regulating Bcl-2/BAX and ERK/Cyclin-D1 in hDPCs. Moreover, synthetic ceramides have potential to stimulate hair growth via Wnt/β-catenin and BMP2/4 signaling in hDPCs. Our results provide novel information that synthetic ceramides such as oleyl and stearyl ceramides might have potential to help hair growth via cell proliferation by regulating gene expression in hDPCs. Although these findings are just laboratory results, our results explained the possibility of stimulating hair growth by treatment with ceramide. Furthermore, our synthetic ceramides might be used as new therapeutic agents to promote hair growth.



 XML Download
XML Download