

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Becker's nevus (BN) was first described by Becker1 in 1949, with cases where unilateral hypertrichosis and hyperpigmentation occurred on upper back. BN clinically shows an irregular well defined brownish macular area, which is frequently associated with hypertrichosis and acneiform lesions. The pathogenesis of BN is not fully understood, and it has been reported to be related with sun-exposure and increased susceptibility and reaction to androgen hormone23.
Melanocytes and peripheral nerve systems are all derived from neural crest, and various pigmentary disorders may accompany abnormalities in neural system or develop within specific dermatome, suggesting that melanocyte and hyperpigmentation may be closely related to neural factors45.
It is known that nerve fibers which secrete neuropeptides are physically connected to melanocytes, and some neuropeptides are responsible for triggering proliferation of melanocytes6. There are also reports that increased number of cutaneous nerve endings have been found in pigmentary disorders such as café au lait macules and congenital giant pigmented nevus78. There are reports of melasma having increased expression of nerve growth factor and neural endopeptidases9.
Several case reports about BN showing linear and segmental configuration suggest a possibility of occurrence of BN in nerve distribution1011. Furthermore, it has been reported recently that protein gene products (PGP) 9.5+ nerve fibers were abundantly distributed in dermis BN compared with normal skin12.
From these perspective that neural factors may be closely related to epidermal pigmentation, we sought to verify factors related to neural process in the lesion of BN including neuropeptides and their receptors and look for possible relationship between these factors and epidermal hyperpigmentation of BN. Among the neuropeptides that can be found in human skin, neuropeptide Y (NPY) is related to neural crest-derived tumors as well as blue, fusiform, halo nevi and melanoma13, and galanin was reported to be expressed in a greater degree in some cutaneous melanomas, which were strongly correlated with α-MSH expression of the same tumors14. Neurotensin (NTS) stimulates human mast cell degranulation and activates mast cells, which is known to contribute epidermal hyperpigmentation in dermatofibroma, urticarial pigmentosa, postinflammatory hyperpigmentation15.
There are no previous reports regarding the role of neuropeptides and nerves in hyperpigmentary lesions of BN. In this study, we investigated whether a linkage may exist among nerve-derived factors such as neuropeptides, distribution of nerve endings, mast cells and hyperpigmentation in BN.
MATERIALS AND METHODS
Patients and samples
Skin specimens used in polymerase chain reaction (PCR) array and real-time PCR were obtained from 3 patients, 41-year-old male, 49-year-old male, and 20-year-old male diagnosed as BN after informed consent of the experiment. The patients had no history of other medical problems. As for immunohistochemical (IHC) study, 3 lesional and 3 normal skin biopsy specimens using a 4-mm disposable punch were used to stain with antibodies associated with NPY, galanin, NTS and their receptors, and PGP 9.5. Normal skin samples were obtained from at least 10 cm from the lesion. This study was approved by Institutional Review Board at Dankook University Hospital (IRB no. 2013-08-002).
PCR array
RT2 Profiler™ PCR Arrays of “Human neurotrophin and receptors” (PAHS-031Z; Qiagen, Valencia, USA) was used in the study. It profiles the expression of 84 genes related to neuronal processes. Genes involved in normal function of the neuronal system and cytokines and receptors involved in neuronal signaling are contained on the array. Skin samples were obtained by biopsy of the lesions using a 4-mm disposable punch from three patients with BN and immediately frozen at −80℃. Total RNA was extracted Using RNeasy mini kit (Cat. No. 74104; Qiagen) from frozen tissue samples. The tissues were ground to powder under frozen conditions and lysed in the lysis buffer provided by Qiagen, Inc. and other steps were performed according to the manufacturer's protocol. Quality and quantity of RNA were determined using the NanoDrop 2000 (Thermo Scientific Inc., Wilmington, DE, USA). The RT reaction was performed with 1 µg total RNA using the RT2 First-Strand Kit (Cat. No. 330401; Qiagen). Randomprimed cDNAs were processed for quantitative real-time reverse-transcriptase PCR of 84 genes involved in neuronal processes and 12 housekeeping genes, including internal controls, by using an RT2 Profiler PCR Array Kit (RT2 Profiler PCR Array Human neurotrophin and receptors, PAHS-031Z; Qiagen) and an Eppendorf RealPlex2 Mastercycler PCR system. PCR products were quantified by measuring SYBR Green (Invitrogen, Carlsbad, CA, USA) fluorescent dye incorporation with ROX dye reference using RT2 SYBR Green ROX qPCR Mastermix (Cat. No. 330623; Qiagen). Using an integrated web-based software package for the PCR Array System, the ΔΔCt-based fold-change calculations from the uploaded raw threshold cycle data were obtained.
Real-time PCR
Real-time PCR arrays were performed by using products of SABiosciences. cDNAs were synthesized using QuantiTect Rev. Transcription Kit (Cat. No. 205311; Qiagen) according to its protocol. The reaction was performed at 42℃ for 15 minutes and then stopped immediately by heating at 95℃ for 5 minutes. Amplification, data acquisition, and the melting curve were carried out by the ABI 7900 Real Time PCR system (Applied Biosystems, Foster City, CA, USA). The PCR cycling program was set as follows: stage 1: 95℃ for 5 minutes, stage 2: 95℃ for 10 seconds followed by 60℃ for 30 seconds repeated for 40 cycles, and stage 3: 95℃ for 15 seconds, 60℃ for 15 seconds and 95℃ for 15 seconds. The cycle threshold (Ct) and melting curve of each gene were automatically established and recorded by the software. The delta Ct (ΔCt) method was used for PCR array data analysis. The primer sequences for real-time PCR is described in Table 1.
Immunohistochemistry
Paraffin-embedded sections (4 µm) of BN lesion and normal skin of the same anatomical site were selected, and IHCl analysis was performed using the commercial kit LSAB®+/horseradish peroxidase (HRP) System (Cat. No. K0679; Dako, Carpinteria, USA). Briefly, after deparaffinization of samples and antigen retrieval, the sections were stained with primary antibodies against NPY (Cat. No. ab112473, 1:50 dilution; Abcam, Novusbio, Santa Cruz, LS Bio, Usbiological, Millipore), NPYR (Cat. No. NBPI-59008, 1:5,000 dilution; Novusbio), NPYR2 (Cat. No. NB100-56480, 1:5,000 dilution; Novusbio) or biotin-conjugated isotype control IgGs and incubated for overnight at 4℃, and antibodies against GAL (Cat. No. sc-166431, 1:2,000 dilution; Santa Cruz), GALR1 (Cat. No. NLS4013, 1:1,000 dilution; Novusbio), GALR2 (Cat no. LS-B4081, 1:500 dilution; LS Bio) were used for staining and incubated for 1 hour at room temperature. NTS (Cat. No. 151977, 1:500 dilution; Usbiological), NTS1R (Cat. No. ab75555, 1:1,000 dilution; Abcam), NTS2R (Cat. No. NLS1269 1:2,000 dilution; Novusbio), PGP 9.5 (Cat. No. AB176-l 1:1,000 dilution; Millipore) Later, the slides were stained with biotinylated anti-rabbit, mouse, and goat immunoglobulin and streptavidin-HRP complex for 30 minutes each and were washed with phosphate-buffered saline. After staining with AEC (Peroxidase substrate kit, SK-4200; Vector Laboratories, Burlingame, CA, USA) as a substrate and dying with Mayer's hematoxylin for 2 minutes, the slides were covered with cover glass and then examined by using light microscope.
Statistical analysis
A paired t-test was employed to determine differences in the expression levels of neuropeptides between lesional and normal skin. Data were valued with in a confidence interval of 95%. A p-value of less than 0.05 was considered statistically significantly different (p<0.05). A p-value of less than 0.1 was considered marginally statistically significantly different (p<0.1).
RESULTS
There were increased level of certain neuropeptides' mRNA expression in lesional skin on PCR array
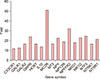
Out of total 84 gene symbols, 34 genes were increased by 3-fold or more in lesional skin compared to normal skin (Table 2). Among them, 16 genes with 10-fold or more increase are as follows by order of fold increases (Fig. 1). Although there are many genes which have a link to melanogenesis through literature research like MC2R, we chose 3 easily approachable genes in our laboratory among those 16 genes.
Significant difference in level of NPY, galanin, NTS and their receptors' mRNA expression between lesional and normal skin was confirmed by real-time PCR
Upon comparing the level between lesional and normal skin, the mRNA levels of NPY, NPY1R and NPY2R were increased by 1.6, 1.4, and 1.5 fold, respectively in lesional skin (Fig. 2A). The mRNA levels of galanin, GAL1R and GAL2R were increased by 1.0, 1.5, and 2.1 fold, respectively in lesional skin when compared to normal skin (Fig. 2B). And, the mRNA levels of NTS, NTS1R and NTS2R were increased by 1.0, 1.2, and 1.2 fold respectively in lesional skin compared to normal skin (Fig. 2C).
In general, statistically significant increase of NPY1R (p<0.05) and marginally significant increase of NPY2R, GAL2R, and NTS2R (p=0.061, p=0.089, p=0.066) was verified in lesional skin compared to normal skin.
Distribution of neuropeptides and their receptors on lesional and normal skin showed difference in immunohistochemistry
NPY, NPY1R and NPY2R were higher degree of expression in epidermis and dermis of lesional skin (Fig. 3E~G) comparing to normal skin (Fig. 3A~C), with NPY being stained in the spinous layer (Fig. 3E), NPY1R being stained throughout the whole layer of epidermis (Fig. 3F) while NPY2R being stained in the basal layer of epidermis (Fig. 3G). Galanin, GAL1R and GAL2R were all expressed in epidermis and dermis of lesional (Fig. 4E~G) and normal skin (Fig. 4A~C), with similar degree of expression when compared to normal skin. GAL and GAL 1R were expressed in whole layer of epidermis (Fig. 4E, F), and GAL2R in spinous layer (Fig. 4G).
NTS and NTS1R were expressed in epidermis and dermis of lesional (Fig. 5E, F), and normal skin (Fig. 5A, B), similar degree of expression when compared to normal skin. NTS2R was expressed with higher degree of expression in lesional skin (Fig. 5G) when compared to normal skin (Fig. 5C). NTS and its receptors were expressed in whole layer of epidermis.
Distribution of PGP 9.5 immunoreactive nerve fibers on lesional and normal skin showed difference in immunohistochemistry
PGP 9.5 immunoreactive nerve fibers were found in the epidermis and dermis, revealing the increased innervations in lesional skin when compared to normal skin. Linear nerve fibers were increased in lesional epidermis (Fig. 6A, B).
Increased mast cells in lesional skin
By toluidine blue stain, an increased number of mast cells was found in perivascular area of lesional skin when compared to normal skin (Fig. 7).
DISCUSSION
The epidermal changes of BN are variable, although it usually shows hyperkeratosis, subtle acanthosis, papillomatosis, flattened rete ridge elongation and basal layer hyperpigmentation. The hyperpigmentation of BN can be treated with various kinds of laser therapy targeting melanocyte and/or melanin such as Q-switched ruby (694 nm), alexandrite and frequency-doubled Nd:YAG (532 nm), however, recurrence is common1617. More invasive treatment include mechanical abrasion, ablative laser treatment, cryotherapy, surgery and these therapies have been either unsuccessful or have resulted in scarring. However, the mechanism behind the pathogenesis of the epidermal hyperpigmentation and the treatment failure of BN remains to be clarified.
It has been linked various pigmentary disorders with neuronal process such as neurofibromatosis, melanocytic nevi, melasma, vitiligo, etc.918192021,22. Several cases about BN showing linear and segmental configuration suggest a possibility of occurrence of BN in nerve distribution1011. Also, there are reports of BN in a patient with neurofibromatosis2324. Recently, Sheng et al.12 reported that PGP 9.5+ nerve fibers were abundantly distributed in dermis. They found that dermal nerve fiber length was higher in the nerve plexus and arrector pili muscles and around eccrine ducts and that dermal nerve fiber length was higher in lesional and perilesional skin compared with normal skin.
Collectively, these previous reports suggest that nerve or neural factors might influence the epidermal hyperpigmentation of BN. However, there are no previous reports about the neuropeptides and nerve in epidermal hyperpigmentary lesions of BN. Therefore, we hypothesized that a linkage may exist between nerve-derived factors such as neuropeptides and melanocytes, and that they may play an important role in the mechanism of hyperpigmentation in BN or recurrence after melanocyte-targeting laser therapy or ablative treatment.
Using lesional and normal tissue specimen from patients with BN, we performed a PCR array to figure out increased neural components, and have found that several neuropeptides including NPY, galanin, NTS and its receptors were significantly increased in lesional skin of BN when compared to normal skin.
NPY is a 36 amino-acid long peptide characterized by many tyrosine residues25. It is mainly produced by the neurons of different brain locations and is stored and released in sympathetic peripheral nerve endings regulating various organ systems26. Other than physiologic functions, NPY can also act as growth and angiogenic factor and are related to neural crest-derived tumors as well as blue, fusiform, halo nevi and melanoma. It has been reported that NPY mediates invasiveness of tumors by modulating angiogenesis, apoptosis, cell adhesion and motility. Furthermore, a positive relationship between the expression of α-MSH and NPY was found in melanocytic lesions13.
NPY was also reported to regulate the secretory activity of melanin-containing cells in Xenopus laevis27. From the reports we can predict that neuropeptides may not regulate synthesis of melanin within melanocytes but rather affect melanosomal transfer to surrounding keratinocytes. It is known in previous reports that NPY is known to be one of the major neuronal peptides found in human skin28. The presence of NPY receptor that couples to inhibitory G protein was shown in keratinocytes, and the activity of the receptor was demonstrated to be downregulated by glucocorticoid treatment in keratinocytes29.
Our study revealed statistically significant increase of NPY1R (p<0.05) and marginally significant increase of NPY2R (p=0.061) level in lesional skin compared to normal skin through real-time PCR. Furthermore, immunohistochemistry showed that NPY, NPY1R, and NPY2R were expressed with higher degree of expression in lesional skin when compared to normal skin. From the fact that NPY, NPY1R and NPY2R expression were significantly increased in lesional skin, it is possible that NPY and its receptors may be involved in epidermal hyperpigmentation of BN by interacting with hormones such as α-MSH or stimulating melanin transfer.
Galanin is a neuropeptide with wide-range of effects within endocrine and nervous system. It was reported that galanin is expressed in a greater degree in some cutaneous melanomas, which were strongly correlated with α-MSH expression of the same tumors14. It appears that galanin expression in pituitary adenomas is linked to expression of ACTH which is a neuropeptide derived from proopiomelanocortin like α-MSH3031. Previous reports revealed that galanin is expressed in keratinocytes, eccrine glands, and around vessels of human skin, suggesting its role other than those attributed to the nervous system32. Interestingly in one study, galanin mRNA level was downregulated in melanocytes after being treated with Q-switched Ruby laser, suggesting that galanin is expressed in human melanocytes33. Furthermore, through the report of expression of both galanin and α-MSH in melanoma, the interaction between galanin and α-MSH may contribute to proliferation of melanocyte14. Other studies reported that galanin receptor mRNA expression was found in both cultured primary and transformed keratinocytes34.
Real-time PCR showed marginally significant increase of GAL2R level in lesional skin compared to normal skin (p=0.089). Galanin, GAL1R and GAL2R were all expressed in lesional (Fig. 5E~G) with similar degree of expression when compared to normal skin in IHC stain. Since the effect of galanin and its receptors in human skin is not well established so far, their role in pathogenesis and recurrence of BN is not clear. According to one report, it has been told that increased levels of galanin-like immunoreactivity and galanin mRNA could be found in cells of inflamed dermis and epidermis, and that galanin or galanin-like peptide is related in inflammatory response35.
Consistently, our study showed that galanin and its receptors were expressed in epidermis and dermis of, and from these findings, it can be inferred that inflammatory process after laser therapy may increase the expression of galanin and/or its receptor resulting the melanocyte proliferation via interaction with α-MSH.
NTS is a single polypeptide chain containing 13 amino acid residues, which works as a neuromodulator in the central nervous system, and as an endocrine agent in the periphery3637. NTS1R is commonly high expressed in melanoma cells, but low expressed in normal immortalized human keratinocyte line HaCaT. SR48692, NTSR1 inhibitor, not only reduced cell proliferation and self-renewal potential in vitro, but also inhibited the tumor growth derived from A375 melanoma cells in NOD/SCID mice in vivo. The peptides corticotropin-releasing hormone (CRH) and NTS have synergistic action stimulating mast cells and NTS triggered human mast cell degranulation and significantly augmented CRH-induced vascular endothelial growth factor release3839. Real-time PCR showed that NTS2R were verified marginally significant increase of NTS2R (p=0.066) in lesional skin compared to normal skin. Immunohistochemistry showed consistently that NTS2R was expressed with higher degree of expression in lesional skin when compared to normal skin. Toluidine blue stain revealed an increased number of mast cells in perivascular area of lesional skin when compared to normal skin. These data support that NTS and its receptors might contribute to melanocyte proliferation and melanogenesis in BN directly or indirectly by stimulating mast cell degranulation.
Furthermore, immunochemical stain revealed that PGP 9.5 immunoreactive nerve fibers were found in the epidermis and dermis, revealing the increased innervations in lesional skin when compared to normal skin. Combined with the previous report showing PGP 9.5+ nerve fibers were abundantly distributed in dermis of BN and dermal nerve fiber length was higher in lesional and perilesional skin compared with normal skin, increased innervation also might be an important to the way in which we understand the pathogenesis of BN.
From the above mentioned role of each neuropeptides, we may hypothesize the way they could be related to hyperpigmentation of BN. Firstly, they may have an effect on hyperpigmentation along with the interaction of increase in α-MSH level. It is well established that elevation in MSH levels may potentiate tyrosinase activity and stimulate melanogenesis of human skin40. Therefore, since high expression of NPY and galanin is related to the expression of α-MSH, it may ultimately result in hyperpigmentation in BN lesions through a set of responses mediated by α-MSH or together with neuropeptides. Secondly, the pigmentation may be caused by inflammatory sequences. NPY and galanin are reported to have role in inflammation, for example, NPY stimulates mast cell degranulation and histamine-release41. It has been reported that inflammatory mediators may increase activity of tyrosinase within melanocytes, which increases melanin production and transfer of melanosomes to keratinocytes42. In general, neuropeptides play an important role in regulating the synthesis and secretion of cytokines including interleukin (IL)-1, IL-2, IL-4, IL-6, and tumor necrosis factor-α4344.
Thus it is possible to speculate that neuropeptides are capable of modulating and interacting with melanocyte immune function during cutaneous inflammation. The limitation of our study includes the fact that the experiment included small number of samples, and the study did not clarify the functional aspect of NPY, galanin, and NTS including their role and effect on proliferation, migration, survival and melanogenesis of melanocytes. Our study was focused on comparing the expression level of genes related to neuronal process such as neuropeptides in both lesional and normal skin of patients with BN, confirming that the expression of neuropeptides including NPY, galanin, and NPY are increased in BN. This result has an important value since it provides a clue that neurologic association should be considered when studying the pathogenesis of epidermal hyperpigmentary disorders. Further studies may be required for precise investigation on roles of these neuropeptides and search for other possible mediators in the pathogenesis of BN.
In conclusion, we hereby propose that NPY, galanin, NTS, their receptors and increased innervation may play a role in the pathogenesis of BN. Furthermore, these neuropeptides and increased innervation might be the possible factors inducing recurrence of BN treated by epidermal ablation and melanocyte or melanin targeted laser therapy. There are many limitations, but we think the results of this study will be valuable as a preliminary data associated with BN and neuropeptides.







 XML Download
XML Download