

 PDF
PDF Citation
Citation Print
Print



Abbreviations
ANRIL
antisense non-coding RNA in the INK4 locus
AS
antisense
c-CBL
casitas B-lineage lymphoma
Cox2
cyclooxygenase 2
DCs
dendritic cells
DM
dermatomyositis
DNA PKcs
DNA-dependent protein kinase catalytic subunit
EGFR
epidermal growth factor receptor
ESR
erythrocyte sedimentation rates
EZH2
enhancer of zeste homolog 2
FAS-AS1
Fas-antisense 1
FLS
fibroblast-like synoviocytes
GAPLINC
gastric adenocarcinoma predictive long intergenic noncoding RNA
Gas5
growth arrest-specific transcript 5
HCC
hepatocellular carcinoma
hnRNP
heterogeneous nuclear ribonucleoprotein
HOTAIR
HOX transcript antisense RNA
ICAM-1
intercellular adhesion molecule 1
KCNQ1OT1
KCNQ1 opposite strand/antisense transcript 1
LCR
locus control region
lincRNA
intergenic long noncoding RNA
lincRNA-EPS
lincRNA erythroid prosurvival
lncRNAs
long noncoding RNAs
miRNAs
microRNAs
MMP
matrix metalloproteinase
Morrbid
myeloid RNA regulator of Bim-induced death
MTX
methotrexate
ncRNA
non-protein coding RNA
NEAT1
nuclear enriched abundant transcript 1
NeST
nettoie Salmonella pas Theiler's
NFAT
nuclear factor of activated T cells
NRON
noncoding repressor of nuclear factor of activated T cells
nt
nucleotides
PRC2
polycomb repressive complex 2
RA
rheumatoid arthritis
PACER
p50-associated COX-2 extragenic RNA
RBM5
RNA-binding motif protein 5
PM
polymyositis
SLE
systemic lupus erythematosus
SLEDAI
systemic lupus erythematosus disease activity index
SS
Sjögren's syndrome
SRP
signal recognition particle
THRIL
tumor necrosis factor-α and heterogenous nuclear ribonucleoprotein L related immunoregulatory lincRNA
Tmevpg1
Theiler's murine encephalomyelitis virus persistence candidate gene 1
mcircRasGEF1B
mouse circRasGEF1B
WDR5
WD40 repeat protein 5
INTRODUCTION
Large portions of the human genome, which had previously been considered as ‘transcriptional noise’ with no coding capacity as protein-coding genes, are estimated to be less than 2% of whole genome (1). The ENCyclopedia of DNA Element Consortium reported that most of the genome is transcribed as non-protein coding RNA (ncRNA) (2). Many recent studies have revealed that ncRNAs are important regulators of gene expression and also play important roles in the pathogenesis of various diseases. ncRNAs are largely categorized into small ncRNAs (<200 nucleotides [nt]) and long ncRNAs (lncRNAs) (>200 nt) according to their transcript size (3). A great deal of research has been focused on microRNAs (miRNAs), one type of small ncRNA. miRNAs have been reported to be involved in the regulation of gene expression and development of autoimmune diseases including systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), systemic sclerosis, Sjögren's syndrome (SS), multiple sclerosis, and type 1 diabetes mellitus (45).
Recently, researchers have started to pay more attention to lncRNAs, which account for major portion of ncRNAs. Compared with miRNAs, the mechanisms of lncRNA function are more diverse due to structural complexity of lncRNAs (36). Early studies primarily showed that lncRNAs are related to diverse cellular responses, including cell differentiation, cell proliferation, and apoptosis (789). Cancer, along with cardiovascular and neurological diseases, was the main focus of lncRNA research (10). However, more recently, accumulating evidence that lncRNAs participate in immune cell differentiation and immune responses, suggests that lncRNAs are essential components in the development of autoimmune diseases (1112131415). In this review, we are going to introduce classification, and mechanism of action of lncRNAs, the influences of lncRNAs on various immune cells, and the role of lncRNAs in autoimmune diseases such as SLE, RA, polymyositis (PM)/dermatomyositis (DM), and SS.
CATEGORIZATION OF LNCRNAS
The lncRNAs are more categorized according to various genomic architectures as well as functional mechanisms (Fig. 1). Based on location in the genome, lncRNAs can be divided into intergenic lncRNAs (lincRNAs) and intronic lncRNAs (316). LincRNAs are transcribed inter genetically from both DNA strands which do not overlap protein-coding genes. Whereas intronic lncRNAs are transcribed from introns of protein-coding genes in either direction and terminate without overlapping exons. Thus, lincRNAs and intronic lncRNAs could be regulated via different transcription activation mechanisms. In addition, lncRNAs are also classified regarding the product orientation of the DNA strand (1718). Sense lncRNAs are transcribed from the sense strand of protein-coding genes and contain exons that overlap with part of protein-coding genes or cover the entire sequence of a protein-coding gene containing an intron. On the contrary, antisense lncRNAs are transcribed from the antisense strand of the protein-coding genes that overlap with exon or intron regions, which cover the entire protein-coding gene. Interestingly, most of the lncRNAs are discovered from protein-coding genes or the antisense of the protein-coding genes using cap-analysis gene expression and oligo-dT guided reverse transcription, suggesting that they also possess 5′ cap and 3′ poly-A tail like mRNA (19).
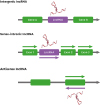 | Figure 1Classification of lncRNAs. The lncRNAs can be classified based upon genomic structures and product orientation of the DNA strand. Based on genomic location, lncRNAs are divided into lincRNAs and intronic lncRNAs. And, according to the direction of transcription, lncRNAs can be classified into sense and antisense lncRNAs.
|
LNCRNAS AS POTENT GENE REGULATORS
Recent studies have shown that lncRNAs are involved in various gene transcription regulation mechanisms through interactions with transcription factors or epigenetic modifiers (Fig. 2). lncRNAs can play as cofactors to modify the activity of transcription factors. For instance, NF-κB-interacting lncRNA binds NF-κB/IκB in a ternary complex, which inhibits IKK-mediated IκB phosphorylation by directly masking the phosphorylation sites (20). lncRNA-Evf2 is transcribed from conserved distal enhancer and recruits the transcription factor DlX2 to the same enhancer to induce expression of adjacent protein-coding genes (21). Some lncRNAs are also involved in post-transcriptional regulation of mRNAs such as capping, splicing, editing, transport, translation, degradation, and stability at various control sites. The lncRNA metastasis-associated lung adenocarcinoma transcript 1 is known to regulate alternate splicing by interacting with multiple splicing factors (22). In addition, lncRNA Gomafu/myocardial infarction-associated transcript is localized to a nuclear domain, which may block spliceosome formation and affect the mRNA splicing by sequestering splicing factor 1 (23).
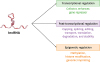 | Figure 2Functions of lncRNAs. lncRNAs regulate gene transcription through interactions with transcription factors or epigenetic modifications. lncRNAs can change activity of transcription factor, or regulate post-transcriptional events of mRNAs, or modify epigenetically.
|
Epigenetic modifications are widely known important gene transcription regulation mechanism. In this context, lncRNAs (antisense non-coding RNA in the INK4 locus [ANRIL], X-inactive specific transcript, HOX transcript antisense RNA [HOTAIR], and KCNQ1 opposite strand/antisense transcript 1 [KCNQ1OT1]) have crucial roles as epigenetic regulators. lncRNA KCNQ1OT1 binds to polycomb repressive complex 2 (PRC2) and the methyl-transferase G9a (EHMT2), whereas ANRIL binds to PRC1 and PRC2 (24). lncRNA HOTAIR forms histone modifying complexes, which coordinates the targeting of specific repressive histone modifying complexes (25). Additionally, lncRNAs have also been proven to X chromosome inactivation, gene silencing, and gene imprinting via regulation of DNA methyltransferases (2426).
REGULATION OF THE IMMUNE SYSTEM BY LNCRNAS
Dysregulation of the immune system can be a major cause of the development of various autoimmune diseases, as well as infectious diseases and cancers. The regulation of gene expression involved in the immune system is particularly crucial for making decisions between immune tolerance and autoimmunity. Recently, increasing evidence has revealed that lncRNAs play an important role in the regulation of the immune system. However, to date, the study of lncRNAs in immune system regulation remains challenging due to the complexity and diversity of the immune system. The immune system is roughly divided into innate and adaptive immunities. Coordination of innate and adaptive immune responses has a critical role in biological and pathological immune responses. Here, we are going to review the roles of lncRNAs in immune responses. Table 1 shows representative lncRNAs that regulate immune responses.
Table 1
Summary of lncRNAs involved in immune responses

| Type of immunity | Source | lncRNAs | Function | Mechanism of action | Reference |
|---|---|---|---|---|---|
| Innate immunity | Myeloid cell | Morrbid | Control lifespan of short-lived myeloid cells (neutrophils, eosinophils, monocytes) | Inhibiting Bcl2l11 (Bim) transcription | (30) |
| DC | lnc-DC | DC differentiation | Phosphorylating STAT3 | (13) | |
| DC macrophage | lincRNA-Cox2 | Regulate expression of proinflammatory genes | Interacting with hnRNP-A/B and hnRNP-A2/B1 | (15) | |
| Macrophage | THRIL | Positively regulate TNF-α expression | Interacting with hnRNPL | (37) | |
| Macrophage | mcircRasGEF1B | Positively regulate ICAM-1 expression | Acting as a miRNA sponge | (38) | |
| Monocyte | PACER | Induce PTGS2 expression | Sequestrating NF-κB subunit p50 | (39) | |
| Monocyte macrophage | lincRNA-EPS | Restrain inflammation | Binding to hnRNPL | (40) | |
| Erythrocyte | |||||
| Macrophage | lnc13 | Restrain inflammation | Binding to hnRNPL | (41) | |
| Fibroblast | Lethe | Restrain inflammation | Binding to NF-κB subunit RelA | (42) | |
| Adaptive immunity | Th1 cell | linc-MAF-4 | Induce Th1 cell differentiation | Suppressing MAF expression | (43) |
| Th2 cell | lincR-Ccr2-5′AS | Induce Th2 cell migration | In a GATA-3 dependent manner | (44) | |
| Th2 cell | TH2-LCR | Positively regulate the transcription of Th2 cytokine gene | Recruiting WDR5-containing complexes | (14) | |
| Th1 cell CD8+ T cell | NeST (Tmevpg1) | Enhance IFN-γ expression | Binding to WDR5 | (47) | |
| CD8+ T cell | lncRNA-CD244 | Suppress TNF-α and IFN-γ expression | Recruiting EZH2 to the Ifng and Tnf promoters | (50) | |
| T cell | NRON | Maintain resting state of T cells | Sequestering phosphorylated NFAT in the cytoplasm | (51) | |
| Treg cell | lnc-EGFR | Stimulate Treg differentiation | Binding to EGFR and inhibiting interaction between EGFR and c-CBL | (53) | |
| B cell | FAS-AS1 | Enhance Fas-mediated apoptosis in B cell lymphoma | Binding to RBM5 and inhibiting RBM5-mediated alternative splicing of FAS pre-mRNA | (54) |
![]()
The role of lncRNAs as regulators of innate immunity
Innate immunity is defined as the first line of host defense against pathogens and induces adaptive immune system to conduct effector functions (2728). The innate immune system is mainly mediated by dendritic cells (DCs), macrophages, and NK cells (29).
There is a lncRNA associated with the regulation of the life span in short-lived myeloid cells. Myeloid RNA regulator of Bim-induced death (Morrbid) is a lncRNA which is highly expressed in short-lived myeloid cells such as neutrophils, eosinophils, and ‘classical’ monocytes (30). These short-lived myeloid cells provide the first line of defense against pathogens and the regulation of their life span is vital for protective host immune responses (3132). Morrbid has been reported to control the lifespan of these cells by repressing the transcription of the pro-apoptotic gene, Bcl2l11, which encodes Bim (30). The expression of Morrbid can be induced by cytokines of the common β-chain receptor family (IL-3, IL-5, and GM-CSF) in vitro (30). Eosinophils from patients with high plasma concentrations of IL-5, who have hypereosinophilic syndrome, show high expression of Morrbid compared with eosinophils from healthy controls (3033). The expression of Morrbid in eosinophils is also positively correlated with plasma IL-5 concentrations (30). The findings suggest that Morrbid might have a significant role in diseases which are related with altered lifespans of short-lived myeloid cells.
DCs are the primary antigen presenting cells for T cells and act as initiators of innate and adaptive immunity (34). Human DCs exclusively express the lncRNA lnc-DC, which was discovered by transcriptome microarray analysis and RNA sequencing (13). When common myeloid progenitor cells or monocytes differentiate into DCs, the expression of lnc-DC is upregulated. lnc-DC knockdown causes considerable change in the regulation of DC function-related genes. Knockdown of lnc-DC diminishes the ability of DCs to uptake antigens and results in the downregulation of molecules associated with T cell activation, including CD40, CD80, CD86, and HLA-DR, and the impairment of CD4+ T cell proliferation and attenuation of IL-12 production upon LPS stimulation (13). lnc-DC, which usually functions in the cytoplasm, interacts with STAT3 required for DC development and function, preventing it from binding to the tyrosine-phosphatase SHP1. Phosphorylation of STAT3 at Tyr705 by lnc-DC leads to translocation of STAT3 to the nucleus, and consequently promotes DC differentiation and activation of the immune system (1335). These data suggest that lnc-DC is crucial for DC differentiation and functions.
There are lncRNAs involved in the induction of inflammation. The expression of lincRNA-cyclooxygenase 2 (Cox2) is markedly upregulated after stimulation with TLR4 agonist in CD11C+ bone marrow-derived human DCs (36). TLR2 ligation induces the expression of lincRNA-Cox2 in murine bone marrow-derived macrophages (15). TLR4 and TLR7/8 ligands also leads to increased expression of both lincRNA-Cox2 and the Ptgs2 (also known as Cox2) gene via the Myd88-NF-kB signaling pathway (15). LincRNA-Cox2 controls the transcription of genes associated with the inflammatory response by interacting with heterogeneous nuclear ribonucleoprotein (hnRNP)-A/B and -A2/B1 (15).
The lncRNA TNF-α and hnRNPL related immunoregulatory lincRNA (THRIL) (also known as linc1992), which is induced by activation of TLR2 signaling in human THP1 macrophages, is a regulator of TNF-α induction. Knockdown of THRIL results in reduced expression of TNF-α mRNA and protein in macrophage cells (37). THRIL binds to hnRNPL and upregulates TNF-α gene transcription (37). Knockdown of Hnrpl leads to a reduction in the levels of TNF-α produced by macrophages.
Mouse circRasGEF1B (mcircRasGEF1B), which is a kind of circular RNA, is expressed in macrophages induced by LPS. LPS-induced expression of mcircRasGEF1B is dependent on NF-κB. The action of mcircRasGEF1B is like a ‘sponge’ for miRNA, which targets intercellular adhesion molecule 1 (ICAM-1) (38). Knockdown of mcircRasGEF1B in LPS-activated macrophages reduces ICAM-1, which is important to initiate inflammation by homing leukocytes to inflammatory sites (38).
The lncRNA p50-associated COX-2 extragenic RNA (PACER) is expressed in human monocytes after stimulation with LPS (39). PACER induces PTGS2, which encodes Cox-2, by sequestrating the NF-κB subunit p50 away from the PTGS2 promoter (39). PACER acts as an activator of the inflammatory response.
There are also lncRNAs associated with restrain inflammatory responses. LincRNA erythroid prosurvival (lincRNA-EPS) is expressed in DCs, macrophages, and erythrocytes. lincRNA-EPS is thought to repress inflammation by binding to hnRNPL. A previous study showed that inflammation was induced in lincRNA-EPS-deficient mice (40).
Similar to lincRNA-EPS, lnc13 functions as a suppressor of inflammation. The expression of lnc13 is observed in macrophages and TLR4 ligation downregulates its expression (41). By binding hnRNPD, lnc13 inhibits immune response genes (41).
The pseudogene lncRNA Lethe, which is named after the ‘river of forgetfulness’ in Greek mythology, is induced in mouse embryonic fibroblasts stimulated by proinflammatory cytokines such as IL-1β and TNF-α (42). Lethe exerts negative regulatory functions upon NF-κB; knockdown of Lethe results in the upregulation of NF-κB targets while overexpression of Lethe decreases the activity of an NF-κB reporter (42). Lethe acts by binding to the NF-κB subunit RelA, inhibiting the formation of active NF-κB complexes. Lethe functions as a restrictor of inflammatory responses.
The role of lncRNAs as regulators of adaptive immunity
Adaptive immunity is highly specific. Pathogen-induced innate immune responses trigger antigen-specific adaptive immune responses. The adaptive immune system mainly consists of T and B cells. Emerging evidence indicates that lncRNAs are associated with adaptive immune responses, although their role is not yet fully understood.
lncRNAs in T cell responses
The lncRNA linc-MAF-4 represses the expression of MAF, a Th2-associated transcription factor and drives T cell differentiation into Th1 cells (42). Downregulation of linc-MAF-4 induces T cell differentiation toward the Th2 cell lineage (43). The linc-MAF-4 recruits chromatin modifiers such as lysine-specific demethylase 1 and enhancer of zeste homolog 2 (EZH2) and inhibits MAF transcription (43). These results suggest that linc-MAF-4 is an important regulator of CD4+ T cell differentiation.
The lncRNA lincR-Ccr2-5′AS is specifically expressed in Th2 cells (44). The lincR-Ccr2-5′AS, which is located at the 5′ end of Ccr2 and expressed via antisense transcription, plays a crucial role in controlling the migration of Th2 cells. GATA-3, the transcription factor of Th2 cells, regulates the expression of lincR-Ccr2-5′AS. The genes, which undergo changes in their expression following lincR-Ccr2-5′AS knockdown, considerably overlap with GATA-3-dependent genes. lincRNA-Ccr2-5′AS is closely related with genes encoding Th2 cell chemokines, including Ccr1, Ccr2, Ccr3, and Ccr5. A previous murine experiment demonstrated that lincR-Ccr2-5′AS-depleted Th2 cells showed impaired migration to lung tissue after in vivo transfer, as compared with lincR-Ccr2-5′AS-sufficient Th2 cells (44).
lncRNA TH2-locus control region (LCR) is located at the 3′ end of the RAD50 gene that encodes a double-strand-break repair protein (45). TH2-LCR positively regulates the transcription of genes that encode Th2 cell cytokines including IL-4, IL-5, and IL-13 (14). The function of TH2-LCR might be associated with the recruitment of the WD40 repeat protein 5 (WDR5)-containing complex to the targeted genomic elements. A previous study has revealed that depletion of TH2-LCR reduces WDR5 recruitment to IL-4 and IL-13, but no changes were found for IL-5 (45).
The lncRNA nettoie Salmonella pas Theiler's (NeST, also known as Theiler's murine encephalomyelitis virus persistence candidate gene 1 [Tmevpg1]) is expressed in Th1 cells, CD8+ T cells, and NK cells (464748). NeST, which is located in the adjacent region to the IFN-γ encoding gene, is the lncRNA associated with T cell activation (49). The expression of NeST depends on the activities of the Th1 transcription factors STAT4 and T-bet, and NF-κB (444748). The expression of NeST leads to the enhancement of IFN-γ production from activated CD8+ T cells. NeST binds to WDR5 and recruits the transcription-activation complex to the Ifng promoter, inducing the transcription of Ifng.
The lncRNA lncRNA-CD244, which functions in CD8+ T cells, is associated with the inhibition of T cell activation (50). CD244, a T cell inhibitory molecule, induces the expression of lncRNA-CD244 and lncRNA-CD244 mediates the suppression of TNF-α and IFN-γ by interacting with EZH2. In contrast, knockdown of lncRNA-CD244 increases the expression TNF-α and IFN-γ and improves CD8+ T cell function.
The lncRNA noncoding repressor of NFAT (NRON) was one of the first lncRNAs discovered in T cells. NRON represses T cell activation by inactivating the calcium-dependent transcription factor, NFAT (51). NFAT exists in a phosphorylated state under steady-state condition, but becomes dephosphorylated when the concentration of intracellular calcium increases (5152). Dephosphorylated NFAT then translocates to the nucleus, inducing T cell activation. NRON sequesters phosphorylated NFAT in the cytoplasm, preventing its translocation to the nucleus. Depletion of NRON, therefore, leads to the nuclear translocation of NFAT, resulting in T cell activation (52).
There is a report that the lncRNA lnc-epidermal growth factor receptor (lnc-EGFR) stimulates Treg differentiation in hepatocellular carcinoma (HCC) tissues (53). Jiang et al. (53) revealed that increased expression of lnc-EGFR in CD4+ T cells was linked with Treg polarization in HCC. lnc-EGFR, which binds to EGFR specifically, blocks EGFR ubiquitination by inhibiting the interaction between EGFR and ubiquitin ligase, casitas B-lineage lymphoma (c-CBL). As EGFR ubiquitination by c-CBL leads to EGFR degradation, blocking EGFR ubiquitination results in maintenance of EGFR activation. So, lnc-EGFR maintains activation of EGFR by inhibiting the interaction between EGFR and c-CBL, leading to the stimulation of Treg differentiation.
lncRNAs in B cell responses
There is a report showing that the lncRNA Fas-antisense 1 (FAS-AS1) regulates B cell function indirectly (54). In human B cell lymphoma, the expression of FAS-AS1 induces Fas-mediated apoptosis. FAS-AS1 binds to RNA-binding motif protein 5 (RBM5) and inhibits RBM5-mediated alternative splicing of FAS pre-mRNA. Alternative splicing of FAS pre-mRNA leads to the inhibition of cell apoptosis.
THE ROLE OF LNCRNAS IN AUTOIMMUNE DISEASES
Autoimmune diseases are thought to be caused by a complex combination of interactions involving genetic, immunological, and environmental factors. In the past, autoimmune diseases were considered to be rare, but now the overall prevalence of autoimmune diseases is reported to be about 3%–5% in the general population, according to improved diagnostic modalities (5556). Despite significant advances in revealing the pathogenesis of autoimmune diseases, the pathogenesis of autoimmune diseases is not yet fully understood. The evidence that lncRNA is involved in immunological responses indicates that the dysregulation of lncRNAs participates in the pathogenesis of autoimmune diseases. Table 2 shows representative lncRNAs that are involved in the development of autoimmune diseases.
Table 2
Summary of lncRNAs involved in autoimmune diseases

| Autoimmune disease | lncRNAs | Source | Clinical application | Reference |
|---|---|---|---|---|
| SLE | NEAT1 | PBMCs | Positively correlate with SLEDAI | (61) |
| Gas5 | Plasma | Negatively correlate with ESR and SLEDAI-2K | (65) | |
| CD4+ T cell | ||||
| B cell | ||||
| RA | HOTAIR | PBMCs | Overexpressed in PBMCs, serum exosome of RA | (71) |
| Serum exosome | Suppressed in differentiated osteoclast and rheumatoid synoviocytes | |||
| Osteoclast | ||||
| Synoviocyte | ||||
| H19 | Synovial tissue | Overexpressed in RA synovial tissue | (72) | |
| lincRNA-p21 | Whole blood | Suppressed in RA | (73) | |
| LOC100652951 | T cell | Overexpressed in RA | (74) | |
| LOC100506036 | ||||
| ENST00000483588 | FLS | Overexpressed in RA | (87) | |
| Positively correlated with C-reactive protein and simplified disease activity index score | ||||
| ENST00000438399 | FLS | Suppressed in RA | (87) | |
| uc004afb.1 | ||||
| ENST00000452247 | ||||
| GAPLINC | FLS | Overexpressed in RA | (75) | |
| DM | ENST00000541196.1 | Muscle | Overexpressed in DM | (81) |
| uc011ihb.2 | ||||
| linc-DGCR6-1 | ||||
| ENST00000551761.1 | Muscle | Decreased in DM | (81) | |
| ENST00000583156.1 | ||||
| SS | Tmevpg1 | CD4+ T cell | Overexpressed in SS | (86) |
| Correlate with SSA, ESR, IgG |
![]()
lncRNAs in SLE
SLE is a systemic autoimmune disease characterized by generation of multiple autoantibodies and immune complex deposition, resulting in damage to organs including the kidneys, lungs, brain, and heart (57). Although the pathogenesis of SLE remains unclear, accumulating evidence suggests that lncRNAs might contribute to its development.
Dysregulated TLR4 signaling have been reported to be associated with the development of SLE (585960). There is a lncRNA, which is associated with SLE by the regulation of TLR4 signaling. A previous study found that the lncRNA nuclear enriched abundant transcript 1 (NEAT1) is associated with the pathogenesis of SLE (61). NEAT1 was upregulated in PBMCs of patients with SLE compared to that in the PBMCs of healthy controls. The expression of NEAT1 was positively correlated with SLE Disease Activity Index (SLEDAI) score which is an index for measuring clinical disease activity of SLE. In vitro study with human monocytic cell line, NEAT1 expression was increased after LPS stimulation. Silencing NEAT1 with siRNA in LPS-stimulated human monocytic cell line led to reduced expression of IL-6, CCL2, and CXCL10, which are reported to be involved in the pathogenesis of SLE. NEAT1 is also involved in the TLR4-mediated inflammatory response by affecting the activation of late mitogen-activated protein kinase signaling pathways.
Recent genome-wide association studies showed that chromosomal region 1q25 is an SLE-susceptible locus (62). The genetic evidence that the lncRNA growth arrest-specific transcript 5 (Gas5) is located within chromosome region 1q25 indicates a possible relationship between Gas5 and SLE susceptibility (63). In the murine model of SLE, Gas5 was linked with increased SLE susceptibility (64). Moreover, in human SLE, the expression of Gas5 in plasma, as well as in CD4+ T cells and B cells, was decreased in patients with SLE compared with healthy controls (65). Furthermore, the plasma expression levels of Gas5 were negatively correlated with erythrocyte sedimentation rates (ESR) and SLEDAI-2K scores in patients with SLE (65).
lncRNAs in RA
RA is a chronic autoimmune disease characterized by synovial inflammation and proliferation, which leads to severe joint destruction. The main clinical symptoms of RA include peripheral joint pain, swelling, and joint deformity, though RA can also affect systemic organs including the lungs, heart, and eyes. RA is a multifactorial disease in which host genetic factors, aberrant immune responses, and environmental factors can all contribute to its development (666768). Recently, several studies have demonstrated that dysregulated lncRNAs play a critical role in the pathogenesis of RA.
One of the first mentioned lncRNAs in RA is HOTAIR. HOTAIR was previously reported as playing an important role in the pathogenesis of cancers (6970). In 2015, Song et al. (71) reported that expression of HOTAIR was upregulated in PBMCs and the serum exosome of patients with RA compared to healthy controls. Moreover, upregulated HOTAIR can promote the migration of active macrophages while downregulated HOTAIR, which was observed in differentiated osteoclasts and rheumatoid synoviocytes, can inhibit the production of matrix metalloproteinase (MMP)-2 and MMP-13. These results indicate that aberrant expression of HOTAIR is involved in the pathogenesis of RA.
H19 was reported to be overexpressed in the synovial tissues of patients with RA compared with those from normal/joint trauma controls (72). Song et al. (71) also reported that high expression of H19 was observed in PBMCs from patients with RA compared to normal PBMCs based on microarray analysis, but microarray analysis data produced from serum exosomes did not support the results. Further studies are therefore needed to determine the role of H19 in the pathogenesis of RA.
A previous study reported that there is a relationship between depressed levels of lincRNA-p21 and increased NF-κB activity in patients with RA (73). In the study, Spurlock et al. (73) demonstrated that the expression of lincRNA-p21 was lower in whole blood from patients with RA, while expression of phosphorylated p65 (RelA), which is a marker of NF-κB activation, was higher compared to that in control subjects. Methotrexate (MTX)-untreated RA patients showed reduced expression levels of lincRNA-p21 and increased levels of p65 compared to MTX-treated RA patients. An in vitro study using primary cells and transformed cell lines showed that MTX upregulated the expression of lincRNA-p21 via DNA-dependent protein kinase catalytic subunit (DNA PKcs). In addition, MTX suppressed NF-κB activity in TNF-α-treated cells in a DNA PKcs-dependent manner, through the induction of lincRNA-p21.
Lu et al. (74) found that lncRNAs, LOC100652951 and LOC100506036, were overexpressed in T cells from patients with RA compared to controls. Use of biological agents reduced the expression levels of LOC100652951 in patients with RA. After activation of Jurkat cells with phorbol 12-myristate 13-acetate and ionomycin, upregulated expression of LOC100506036, but not LOC100652951, was observed. Furthermore, knockdown of LOC100506036 in activated Jurkat cells leads to the downregulation of sphingomyelin phosphodiesterase 1 and NFAT, both of which contribute to inflammatory responses.
There was a report that the expression of lncRNA ENST00000483588 was increased and that the expression of three lncRNAs, including ENST00000438399, uc004afb.1, and ENST00000452247, were decreased in fibroblast-like synoviocytes (FLS) from patients with RA compared to controls (74). Among four lncRNAs, the expression level of ENST00000483588 showed a positive relationship with C-reactive protein and the disease activity index score of RA. The results suggest that those lncRNAs may play a role in the pathogenesis of RA, although further studies are needed to confirm this.
Recently, Mo et al. (75) reported the relationship between lncRNA gastric adenocarcinoma predictive long intergenic noncoding RNA (GAPLINC) and RA. The expression level of GPALINC was higher in FLS from patients with RA than in FLS from patients with traumatic injuries. GPALINC knockdown suppressed proliferation, migration, invasion, and the production of inflammatory cytokines and proteinases in RA FLS. Given that silencing of GAPLINC led to the increased expression of miR-382-5p and miR-575, GAPLINC is thought to play a role as a miRNA sponge. The results suggest the possibility that aberrant regulation of lncRNA GAPLINC contributes to the development of RA.
lncRNAs in PM/DM
PM and DM are idiopathic inflammatory myopathies which share some clinical features and show increased serum levels of muscle enzymes (767778). PM and DM both present with symmetric, proximal muscle weakness and can also present with interstitial lung disease. However, they differ in some clinical aspects such as the presence of skin lesions, the histopathology of affected muscle, and prognosis.
There is a report that PM/DM are related with lncRNA 7SL expression, which is the RNA component of signal recognition particle (SRP) (79). It is well-known that patients with DM or PM can have autoantibodies against SRP (80). In the study, PM/DM patients with anti-SRP antibodies were shown to also possess autoantibodies against 7SL RNA. The presence of autoantibodies against 7SL RNA is also related with ethnic background, clinical manifestations, and seasonal onset of the disease. The study suggests that autoantibodies against 7SL RNA may be used as a serological marker for a subset of PM/DM.
Peng et al. (81) found 1,198 differentially expressed lncRNAs in muscles from patients with DM compared with muscles from healthy controls, using a microarray analysis. Among those lncRNAs, they validated five lncRNAs, including ENST00000541196.1, uc011ihb.2, linc-DGCR6-1, ENST00000551761.1, and ENST00000583156.1. According to their bioinformatics prediction, linc-DGCR6-1 regulates the USP18 gene, which is type 1 interferon-inducible gene and is mainly found in perifascicular areas of muscle fibers in patients with DM. These results indicate that dysregulated lncRNAs can play a role in the pathogenesis of DM.
lncRNAs in SS
SS is a chronic, systemic autoimmune disease characterized by dry eye and dry mouth symptoms resulting from inflammation of exocrine glands (82). In addition to glandular manifestations, patients with SS can have various extraglandular manifestations which include pulmonary, skin, articular, renal, and neurological manifestations (83). The production of autoantibodies against SSA or SSB is one of the hallmarks of SS (84). The pathogenesis of SS is multifactorial and is not yet fully understood.
Among various immunologic factors causing SS, Th1 response has been reported to be play an important role in the development of SS (85). There is a study that lncRNA Tmevpg1, which is known to regulate Th1 responses, is implicated in the pathogenesis of SS. The study suggests that there is a relationship between SS and lncRNA Tmevpg1 (86). Increased expression of Tmevpg1 was observed in CD4+ T cells in patients with SS compared with CD4+ T cells of healthy donors. The study also revealed that the expression levels of Tmevpg1 were correlated with the presence of SSA, as well as the levels of ESR and IgG. This study indicates that lncRNA Tmevpg1 can play a role in the pathogenesis of SS.
CONCLUSION
We have reviewed lncRNAs associated with immune cell functions and the pathogenesis of autoimmune diseases. Accumulating evidence suggests the possibility that lncRNAs may be strong candidates for immune regulators and could also be prime suspects in the development of autoimmune diseases. However, only a few studies have been conducted in autoimmune diseases so far. Therefore, further studies on lncRNAs in autoimmune diseases will be essential to answer questions about their role in the pathogenesis of autoimmune diseases. Moreover, future studies may provide novel molecular targets for accurate diagnosis and treatment of autoimmune diseases.
 XML Download
XML Download