

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atherosclerosis, a chronic multi-factorial vascular disease characterized by chronic inflammation of the arterial wall, is a leading health issue worldwide among older adults.1 Endothelial dysregulation and apoptosis have been considered as the initiating factors in the development and pathogenesis of atherosclerosis.2 Epidemiological studies have suggested that oxidized low-density lipoprotein (ox-LDL), a common factor used to establish experimental models of atherosclerosis, is able to induce apoptosis of endothelial cells (ECs) and has been considered as a critical risk factor for the development of atherosclerosis.3 Therefore, inhibition of ox-LDL-induced EC apoptosis may be utilized as a promising novel therapeutic approach in the treatment of atherosclerosis.4 Although various cardiovascular risk factors have been shown to contribute to endothelial dysregulation and apoptosis, the molecular mechanisms underlying EC apoptosis still remain largely elusive.
In recent years, increasing studies have focused on the roles of microRNAs (miRNAs) in vascular diseases. miRNAs are a group of evolutionarily conserved, small, endogenous, non-coding RNAs of 18–22 nucleotides in length. They negatively regulate gene expression by pairing with the 3′untranslated region (3′UTR) of target mRNA at the post-transcriptional level, resulting in a decrease of protein expression via either mRNA degradation or translation inhibition.5 miRNAs are implicated in diverse pathophysiological processes, including differentiation, growth, proliferation, and apoptosis.6 Recent studies have shown that miRNAs are abnormally expressed at different stages of atherosclerosis.7 Moreover, aberrantly expressed miRNAs have been found to contribute to the development and progression of atherosclerosis via multiple target factors or key pathways.8 miR-590, a miRNA located on human genome chromosome 7q11.23, is downregulated and plays critical roles in cardiovascular diseases, including atherosclerosis.910 Importantly, a previous study has well documented the anti-apoptotic effects of miR-590 on ox-LDL-treated ECs.10 However, the mechanism underlying the anti-apoptotic effects of miR-590 in ox-LDL-treated ECs remains to be further addressed.
In the present study, we revealed that miR-590 is downregulated in apolipoprotein E-deficient (apoE−/−) mice fed a high-fat diet (HFD) and ox-LDL-treated ECs. Moreover, miR-590 overexpression inhibited atherosclerotic lesions in HFD-induced apoE−/− mice and promoted the proliferation and inhibited apoptosis in ox-LDL-treated ECs via inactivation of the toll-like receptor 4 (TLR4)/nuclear factor kappa B (NF-κB) pathway. Therefore, miR-590 may serve as a potential therapeutic strategy for the treatment of atherosclerosis.
MATERIALS AND METHODS
Animal models and quantification of atherosclerotic lesions
The animal experiments were approved by the People's Hospital of Zhengzhou University and performed in accordance with the National Institutes of Health Guidelines on the Use of Laboratory Animals. Male apoE−/− mice of C57BL/6 J background and wild-type C57BL/6 J controls (6-week old; weight, 22±5 g) were obtained from the People's Hospital of Zhengzhou University and housed under sterile animal room conditions under a 12 h light and dark cycle at a controlled temperature of 25℃ with free access to food and water. All apoE−/− mice were randomly allocated to three groups (n=10/group): the control group, miR-control group, and miR-590 group. The apoE−/− mice were fed HFD (10% lard, 10% cholesterol, 2% cholate, and 78% basal feed) for 12 weeks to induce atherosclerosis. The experimental mice were injected with miR-control or miR-590 agomir (miR-590) at a dose of 40 mg/kg once every 4 weeks via the tail vein after starting HFD. After 12 weeks, the mice were sacrificed, and the whole length of each aorta and aortic sinuses were resected and stored at −80℃ for further use.
Histological evaluation of atherosclerotic lesions
Isolated aortic sinuses were immediately fixed with 4% paraformaldehyde, dehydrated, embedded in paraffin, and sectioned into 4-µm serial sections. For atherosclerotic lesion examination in aortic sinuses, serial paraffin sections were stained with hematoxylin and eosin (H&E) (Sigma, St. Louis, MO, USA) and observed using an Olympus fluorescent microscope (Olympus Corp., Tokyo, Japan). To assess atherosclerotic plaque formation in atherosclerotic lesions, aortic sinus sections were immersed in 100% isopropanol and examined using Evans blue (Abcam, Cambridge, MA, USA). Atherosclerotic lesion area was quantified using Image Pro Plus 6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Cell culture and transfection
Primary human aortic endothelial cells (HAECs) were obtained from the American Type Culture Collection (Manassas, VA, USA) and cultured in RPMI-1640 medium (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; GE Healthcare Life Sciences, Logan, UT, USA), 100 U/mL penicillin G, and 100 µg/mL streptomycin at 37℃ in a 5% CO2 atmosphere. miR-590 mimics (miR-590), miRNA negative control (miR-con), miR-590 inhibitors (anti-miR-590), inhibitor negative control (anti-miR-con), siRNA specially against TLR4 (si-TLR4), siRNA negative control (si-con), pcDNA-TLR4 (TLR4), and pcDNA empty vector (pcDNA) were obtained from RiboBio Co., Ltd. (Guangdong, China). When grown to 70–80% confluence, HAECs were transiently transfected with miRNAs (100 nM), siRNAs (100 nM), and pcDNA plasmids (1 µg/mL) using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA). One day after transfection, HAECs were exposed to 100 µg/mL of ox-LDL (UnionBiol, Beijing, China) for a further 24 h.
Cell viability assay
The 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay was conducted to assess cell viability of HAECs. In brief, HAECs were seeded into 96-well plates with 200 µL of culture medium at a density of 2×104 cells per well and transfected with miR-590, miR-con, anti-miR-590, anti-miR-con, si-TLR4, si-con, anti-miR-590+si-TLR4, or anti-miR-con+si-TLR4, or exposed to 50 µM PDTC (Sigma) for 24 h, followed by treatment with 100 µg/mL of ox-LDL for 24 h. Then, 20 µL of MTT solution (5 mg/mL; Sigma) was added to each well and incubated for another 4 h at 37℃. Subsequently, the supernatants were discarded and 150 µL of dimethyl sulfoxide was added to dissolve the formazan crystals. The optical density at 490 nm was measured by a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Flow cytometry analysis
The apoptosis of treated HAECs was assessed using Annexin V/fluorescein isothiocyanate (FITC) and propidium iodide (PI) apoptosis detection kits (BD Biosciences, San Jose, CA, USA). The HAECs treated as stated above in 1 mL of culture medium were collected, washed with PBS twice, digested, and resuspended in 200 µL of binding buffer at a density of 2×105 cells/mL. Then, the cells were double stained with 5 µL of FITC-Annexin V and 5 µL of PI for 15 min in the dark. Finally, the apoptotic percentage of cells was detected using a FACScan flow cytometer (BD Biosciences). TLR4 protein levels on the cell surface were evaluated by flow cytometry. Briefly, cells after different treatments were collected by incubation with trypsin under cell culture condition for 2 minutes. Excess trypsin was deactivated using cell culture media, and the cells were labeled with FITC-conjugated antibody against TLR4 (Abcam). The cells were then subject to flow cytometry to analyze TLR4 protein levels on the cell surface.
RNA extraction and quantitative real-time PCR
Total RNA was extracted from the aorta or treated HAECs using TRIzol reagent (Invitrogen). For the determination of miR-590, total RNA was reversely transcribed into cDNA using the TaqMan MicroRNA Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA). A TaqMan miRNA assay was performed to quantify the expression of miR-590. Quantitative real-time PCR (qRT-PCR) was performed on the CFX96™ Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). GAPDH were used as the internal controls for miR-590 expression.
Western blot
Whole cell extracts were extracted from treated HAECs or the isolated aorta using RIPA lysis buffer (Invitrogen) containing protease inhibitors (Sigma). Equal amounts of proteins were subjected to 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). Subsequent to blocking with 5% nonfat milk in TBS containing 0.05% Tween-20 for 2 h, the membranes were incubated at 4℃ overnight with the primary antibodies, followed by incubation with horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 1 h at room temperature. The immunoblotting signals were visualized using an Enhanced Chemiluminescence Western Blotting Analysis kit (GE Healthcare, Solingen, Germany). Antibodies against Cleaved poly (ADP-ribose) polymerase (PARP) Cleaved Caspase-3, α-smooth muscle actin (α-SMA), and Vimentin were purchased from Cell Signaling Technology (Danvers, MA, USA), and antibodies against TLR4, phosphorylated-p65 (p-p65), p65, p-IκBα, IκBα, and GAPDH were all obtained from Santa Cruz Biotechnology. Pecam-1 antibody was obtained from BD Biosciences.
Luciferase reporter assay
The 3′UTR sequences of TLR4 containing the putative binding sites of miR-590 were synthesized and inserted into pMIR-REPORT luciferase reporter plasmids (Promega, Madison, WI, USA) to produce TLR4-WT. A pMIR-REPORT plasmid containing TLR4 mRNA 3′UTR with a mutant sequence in the miR-590 binding sites were synthesized by Suzhou GenePharma Co., Ltd. (Jiangsu, China) and named as TLR4-MUT. HAECs cells were seeded into 96-well plates with 200 µL of culture medium and cotransfected with 50 nM miR-590, anti-miR-590, or matched controls and 100 ng of the luciferase vectors using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's protocol. The cells were harvested at a point 48-h posttransfection, and relative luciferase activity was detected using a Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
In vitro experiments were performed in triplicates. All results are shown as the mean±standard deviation from three independent experiments. All statistical analyses were performed using SPSS version 11.0 software (SPSS Inc., Chicago, IL, USA) with one-way analysis of variance analysis or Student's t test. p<0.05 was considered statistically significant.
RESULTS
MiR-590 overexpression inhibits atherosclerotic lesions in HFD-induced apoE−/− mice
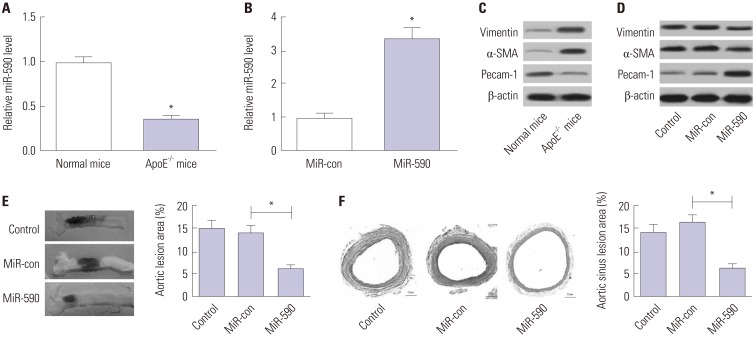
qRT-PCR analysis initially demonstrated that miR-590 expression was abnormally downregulated in the aorta tissue derived from apoE−/− mice, compared with that in normal mice (Fig. 1A). Additionally, miR-590 expression was upregulated after miR-590 injection (Fig. 1B). We found that Pecam-1 levels were reduced and that α-SMA and Vimentin levels were increased in aortic arches from apoE−/− mice in comparison with the normal mice (Fig. 1C), while these changes were significantly reversed following transfection with miR-590 (Fig. 1D). To assess the effects of miR-590 on the development of atherosclerosis in apoE−/− mice, we evaluated atherosclerotic lesions in aortic sinuses by Evans blue and H&E staining. As shown in Fig. 1E and F, miR-590 overexpression significantly suppressed atherosclerotic plaque formation and reduced atherosclerotic lesion area in HFD-induced apoE−/− mice in comparison with the miR-control group, suggesting that forced expression of miR-590 attenuated atherosclerotic lesion in HFD-induced apoE−/− mice.
MiR-590 overexpression promotes proliferation and inhibits apoptosis in ox-LDL-treated HAECs
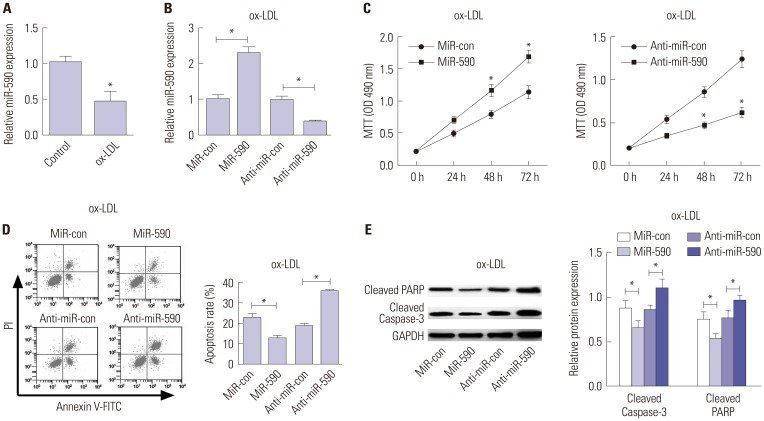
We initially analyzed the expression of miR-590 in HAECs under the condition of ox-LDL administration. qRT-PCR analysis demonstrated that miR-590 expression was significantly decreased in ox-LDL-treated HAECs, compared with control cells (Fig. 2A). To further evaluate the effects of miR-590 on the development of atherosclerosis in vitro, HAECs were transfected with miR-590, anti-miR-590, or matched controls following ox-LDL stimulation for 24 h. As a result, miR-590 transfection significantly increased the expression of miR-590, while anti-miR-590 transfection significantly decreased the expression of miR-590 in ox-LDL-treated HAECs (Fig. 2B). Subsequently, MTT assay revealed that cell proliferation was significantly facilitated by regained expression of miR-590 in HAECs following ox-LDL challenge, compared with the miR-con group (Fig. 2C). On the contrary, inhibition of miR-590 by anti-miR-590 led to a significant inhibition of cell proliferation in ox-LDL-treated HAECs (Fig. 2C). Meanwhile, flow cytometry analysis proved that increasing miR-590 expression significantly hindered apoptosis of ox-LDL-treated HAECs, while anti-miR-590 triggered the opposite effects (Fig. 2D). Consistent with the flow cytometry analysis, Western blot analysis demonstrated that cleavage of PARP and Caspase-3 was significantly attenuated by miR-590 overexpression, but was enhanced following the transfection of anti-miR-590 in ox-LDL-treated HAECs (Fig. 2E). These above results indicated that miR-590 overexpression promotes proliferation and inhibits apoptosis in ox-LDL-treated HAECs.
TLR4 is a direct target of miR-590 in ox-LDL-treated HAECs
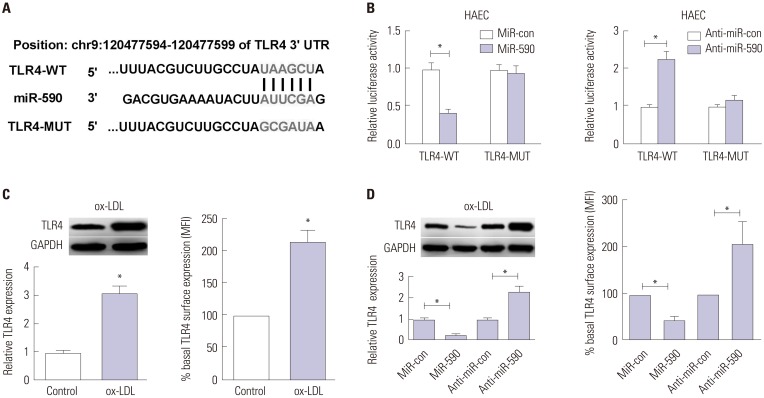
To explore the molecular mechanism by which miR-590 regulates the proliferation and apoptosis of ox-LDL-treated HAECs, bioinformatics analyses were employed to predict the potential targets of miR-590. TLR4, which was reported to play a crucial role in the development of atherosclerosis,11 was identified as a potential target of miR-590. To verify this prediction, the WT or MUT 3′UTR sequences of TLR4 containing the putative binding sites of miR-590 were inserted into pMIR-REPORT plasmids, as shown in Fig. 3A. The results from luciferase reporter assay demonstrated that miR-590 overexpression resulted in a marked decrease of luciferase activity of the 3′UTR of TLR4, but not that of the TLR4-MUT in HAECs cells (Fig. 3B). In contrast, anti-miR-590 led to an obvious elevation of luciferase activity of TLR4-WT, but had no influence on the luciferase activity of TLR4-MUT (Fig. 3B). Additionally, we found that both cellular and cell surface levels of TLR4 protein were significantly increased in ox-LDL-treated HAECs versus their respective controls (Fig. 3C). Moreover, re-expression of miR-590 notably decreased TLR4 protein levels, and miR-590 inhibition distinctly increased TLR4 protein level in ox-LDL-treated HAECs (Fig. 3D). These results confirmed the authentic binding between miR-590 and TLR4.
MiR-590 overexpression antagonizes the apoptosis-promoting role of TLR4 in ox-LDL-treated HAECs
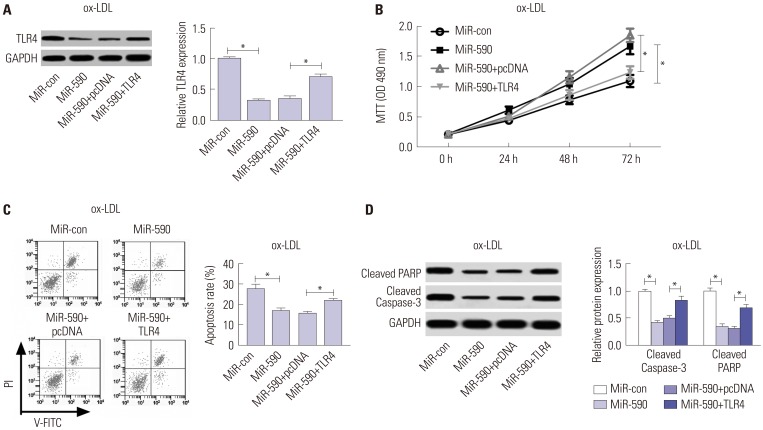
To further address whether miR-590s could attenuate the apoptosis of ox-LDL induced HAECs by targeting TLR4, HAECs were transfected with TLR4 overexpressing plasmid to restore TLR4 protein levels in miR-590 overexpressing HAECs under ox-LDL treatment. Western blot results indicated that TLR4 protein levels in HAEC under ox-LDL treatment could be significantly reduced by miR-590 overexpressing, which was largely restored by TLR4 overexpressing plasmid transfection (Fig. 4A). MTT assay results demonstrated that cell proliferation of ox-LDL-treated HAECs was significantly increased by miR-590 overexpression, which was largely abolished by TLR4 restoration (Fig. 4B). Furthermore, flow cytometry analysis results showed that miR-590 overexpression significantly attenuated ox-LDL induced apoptosis in HAECs, which was weakened by TLR4 restoration (Fig. 4C). The influence of miR-590 overexpression and TLR4 restoration on ox-LDL induced HAECs apoptosis was also verified by Western blot analysis of PARP protein cleavage and caspase-3 activation (Fig. 4D). These results indicated that miR-590 could attenuate ox-LDL induced cell apoptosis in HAECs by targeting TLR4.
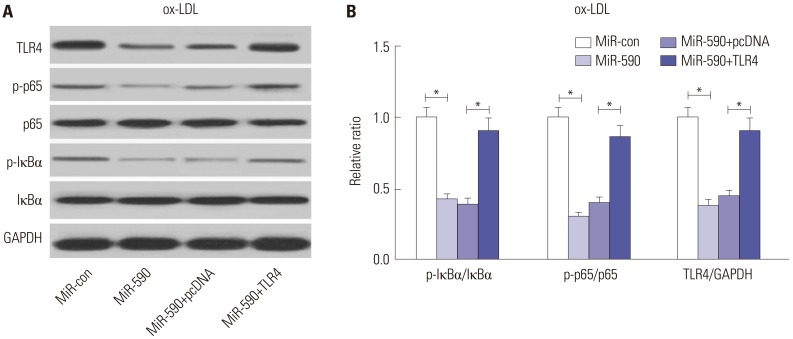
MiR-590 overexpression suppresses the TLR4/NF-κB pathway in ox-LDL-treated HAECs
Since the TLR4/NF-κB pathway has attracted great attention due to its crucial role in atherosclerosis,12 we further analyzed the effects of miR-590 on the TLR4/NF-κB pathway. Western blot analyses revealed that miR-590 overexpression specifically suppressed the protein levels of TLR4, p-IκBα/IκBα ratio, and p-p65/p65 ratio, which was partially recuperated by forced expression of TLR4 (Fig. 5A and B) in ox-LDL-treated HAECs, suggesting that ectopic expression of miR-590 inhibited the activation of the TLR4/NF-κB pathway in ox-LDL-treated HAECs.
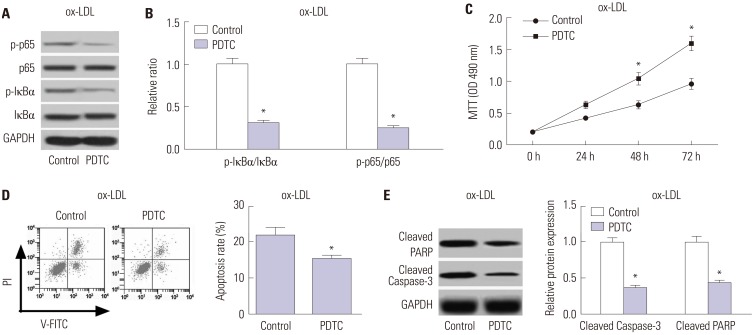
Inhibition of the TLR4/NF-κB pathway promotes cell proliferation and impedes apoptosis in ox-LDL-treated HAECs
PDTC, a NF-κB inhibitor, was used to suppress the TLR4/NF-κB pathway. As shown in Fig. 6A and B, PDTC treatment triggered a substantial decline of the ratio of p-IκBα/IκBα and p-p65/p65 in ox-LDL-treated HAECs, suggesting the suppression of the TLR4/NF-κB pathway by PDTC. Additionally, PDTC exposure dramatically promoted cell proliferation in ox-LDL-treated HAECs, compared with the control group (Fig. 6C). Moreover, PDTC treatment exerted a significant inhibitory role in apoptosis in ox-LDL-treated HAECs relative to the control group (Fig. 6D), as demonstrated by flow cytometry analysis. Furthermore, PDTC also greatly diminished the protein levels of Cleaved PARP and Cleaved Caspase-3 in ox-LDL-treated HAECs with respect to the control group (Fig. 6E). Together, these results demonstrated that inhibition of the TLR4/NF-κB pathway facilitated cell proliferation and restrained apoptosis in ox-LDL-treated HAECs.
DISCUSSION
Currently, substantive studies have revealed that the progression of atherosclerosis is a complex, multiple-step process.13 EC dysfunction, which may be affected by EC proliferation and apoptosis, is regarded as a major contributor to the pathogenesis of atherosclerosis.14 Up to now, the precise mechanism underlying the pathogenesis of atherosclerosis still remains largely unknown. In the present study, we demonstrated the anti-apoptotic effects of miR-590 in ox-LDL-treated HAECs. Mechanistically, the anti-atherosclerotic effect of miR-590 was mediated through inhibition of the TLR4/NF-κB pathway. Therefore, miR-590 may be a novel target for the prevention and treatment of atherosclerosis.
Recently, miRNAs are attracting more attention due to their important regulatory roles in the progression of atherosclerosis.15 For example, miR-98 was reported to promote the proliferation and alleviate apoptosis of human umbilical vein ECs (HUVECs) exposed to ox-LDL.16 Additionally, it was demonstrated that miR-126 alleviated ox-LDL-induced HUVECs injury through restoring autophagy flux via repressing the PI3K/Akt/mTOR pathway.17 Moreover, it was revealed that downregulation of miR-497 suppresses ox-LDL-induced lipid accumulation in THP-1 macrophages via targeting of apelin and thus represented a potential therapeutic target for atherosclerosis.18 Of note, it was previously demonstrated that miR-590 is downregulated in Angiotensin (Ang) II-induced HUVECs and that overexpression of miR-590-5p attenuates Ang II-induced EC apoptosis and decreases reactive oxygen species (ROS) generation by downregulating low-density lipoprotein receptor 1 (LOX-1).19 Moreover, miR-590 agomir could attenuate lipid accumulation and proinflammatory cytokine secretion and could prevent atherosclerosis in apoE−/− mice.2021 In accordance with these previous studies, we provided evidence indicating that miR-590 is downregulated in the aorta tissues from HFD-fed apoE−/− mice and ox-LDL-treated HAECs. Specifically, exogenous overexpression of miR-590 attenuated atherosclerotic lesions in HFD-fed apoE−/− mice, preserved cell proliferation, and suppressed apoptosis in ox-LDL-treated HAECs, suggesting that miR-590 exhibits anti-atherosclerotic activity.
TLRs are evolutionarily preserved pattern recognition receptors and have recently attracted increasing attention due to its link between innate immunity, inflammation, and atherosclerosis.2223 As the most widely studied receptor in the TLR family, TLR4 is abundantly expressed in atherosclerotic lesions at the different stages of atherosclerosis, including macrophages and ECs within the lesion.24 It was reported that TLR4 deficiency could attenuate aortic atherosclerotic lesion areas and inflammatory cytokine levels in apoE−/− mice, suggesting a crucial role of TLR4 in the pathogenesis of atherosclerosis.25 NF-κB is an important downstream mediator of the TLR4 signaling pathway.26 TLR4 can induce activation of NF-κB linked to the transcription of the expressions of many proinflammatory-related genes, resulting in increased endothelial injury and lipid deposition and thereby contributing to the formation and development of atherosclerosis.27 A previous study demonstrated that the TLR4/NF-κB pathway contributes to chronic unpredictable mild stress-induced atherosclerosis through activation of proinflammatory cytokines in apoE−/− mice.28 In the present study, we deemed that TLR4 is a direct target of miR-590 and that miR-590 negatively regulates TLR4 expression in ox-LDL-treated HAECs. TLR4 was found to be upregulated in ox-LDL-treated HAECs. Rescue experiments demonstrated that anti-miR-590 reversed TLR4 knockdown-mediated promotion of cell proliferation and suppression of apoptosis in ox-LDL-treated HAECs, suggesting that miR-590 regulates the proliferation and apoptosis of ox-LDL-treated HAECs by targeting TLR4. Mechanistically, we further discovered that miR-590 inhibits activation of the TLR4/NF-κB pathway in ox-LDL-treated HAECs and that inhibition of the TLR4/NF-κB pathway by PDTC promotes cell proliferation and impedes apoptosis in ox-LDL-treated HAECs, indicating that miR-590 rescues proliferation and blocks ox-LDL-induced apoptosis in HAECs via inactivation of the TLR4/NF-κB pathway. It was previously reported that the mechanism underlying the anti-apoptotic effects of miR-590 in ox-LDL-treated ECs was involved, in part, in the LOX-1-ROS-p38MAPK-NF-κB signaling cascade and p53-Bcl-2/Bax-caspase-3 signaling pathway.10 Antagonizing the TLR-4/NF-κB signaling pathway has been well demonstrated to ameliorate atherosclerosis.293031 A possible ligand for TLR-4 that is involved in atherosclerosis development is high-mobility group protein 1 (HMGB1), which is often released by apoptotic cells, leaked from necrotic/necroptotic cells, or secreted by inflammatory signal-stimulated cells into extracellular space. The pro-atherogenic role of HMGB1 has been summarized by Zhou, et al.32 in a short letter. Given that ox-LDL could induce HMGB1 release from in vitro cultured ECs, we speculated that HMGB1 must be involved in ox-LDL-induced EC apoptosis in vitro and atherogenesis in vivo in such a miR-590/TLR-4-regulated way. This hypothesis will be investigated in our future work.
TLR4 expression was also found to be regulated by other miRNA in atherosclerotic cells, such as miR-20a33 in ECs and miR-181a, miR-223 and miR-155 in macrophages.343536 Besides ApoE knockout mice, several other mouse models are available to establish atherosclerosis models in vivo, such as low-density lipoprotein receptor (LDL-r) knockout mouse.37 In this research, we only used ApoE knockout mice for the in vivo experiments, although we speculated that using LDL-r knockout mice should yield similar results. The major cause of atherogenesis in ApoE knockout mice and LDL-r knockout mice is hypercholesterolemia; the difference is that this pathologic condition in ApoE knockout mice is developed spontaneously, while LDL-r knockout mice need excess cholesterol intake from diet to develop hypercholesterolemia and atherosclerosis. Never the less, it might worth to verify whether miR-590 and TLR4 play similar roles in atherogenesis in LDL-r knockout mice as what we have revealed in ApoE knockout mice.
In summary, the present study demonstrated that miR-590 is downregulated in the aorta tissues from HFD-fed apoE−/− mice and ox-LDL-treated HAECs. Moreover, miR-590 overexpression attenuated atherosclerotic lesion in HFD-fed apoE−/− mice and promoted proliferation and blocked ox-LDL-induced apoptosis in HAECs through inhibition of the TLR4/NF-κB pathway, providing new insights into the molecular mechanism underlying the pathogenesis of atherosclerosis. Therefore, miR-590 may serve as a potential therapeutic target for atherosclerosis when it is overexpressed in ECs.
 XML Download
XML Download