

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Tobacco smoking has a documented impact on human health, and in recent years many studies have found that smoking is closely associated with high caries incidence [123]. Nicotine, the primary bioactive and addictive agent in tobacco, and its metabolites are found in blood, saliva, urine, and breast milk of smokers [456]. An increased nicotine load is observed in the oral cavity of smokers [7], and the concentration of this load ranges from 0 to 2.27 mg/mL in saliva [789]. The nicotine level and its metabolites in the oral fluid are found to be 100-fold greater compared to blood. The presence of some nicotine in breast milk has been reported, which suggests that breastfed infants might be exposed to nicotine [45].
Several in vitro studies reported that nicotine upregulates the metabolic activity and enhances the growth of Streptococcus mutans (S. mutans). In 2012, Huang et al. [10] investigated the effects of nicotine on seven S. mutans strains and demonstrated that 0.25–8 mg/mL of nicotine increases S. mutans biofilm formation significantly while higher concentrations killed the bacteria. In addition, they found that the minimum inhibitory and minimum bactericidal concentrations (MIC and MBC, respectively) for nicotine were 16 and 32 mg/mL, respectively. The same conclusion was reported by Li et al. [11] who concluded that nicotine could increase the growth of S. mutans over Streptococcus sanguinis in dual species experiments. It was reported that nicotine was able to increase the number of S. mutans and the amount of extracellular polysaccharide (EPS) synthesis, the main component of the biofilm matrix (Figure 1A). Nicotine increases glucan-binding protein A (GbpA) expression up to 4.0-fold and glucosyltransferase (Gtfs) expression up to 2.2-fold [12]. A recent metabolomic study indicated that nicotine increases lactate concentration of S. mutans by 2-fold [13]. Nicotine also increases the biofilm formation of Scardovia wiggsiae, a Gram-positive bacillus that is associated with early childhood caries [14]. In addition, Tanaka et al. [15] suggested that smoking during pregnancy or in the postnatal period could increase the risk of caries in the primary dentition of the infants. Moreover, nicotine increases the growth of many other oral species such as Prevotella gingivalis, Lactobacillus casei, Actinomyces viscosus, Rothia dentocariosa, Enterococcus faecalis, and Candida albicans [16].
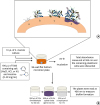 | Figure 1(A) Stages of biofilm formation: (a) Pellicle formation; (b) Binding of single organisms and multiplication; (c) Continued growth; (d) Biofilm maturation that triggers surface demineralization. (B) Schematic diagram of initiation of the biofilm using the 96-well microtiter plate crystal violet staining assay.TSBS, tryptic soy broth (TSB) supplemented with 1% sucrose; NaCl, sodium chloride; KCl, potassium chloride; KI, potassium iodide.
|
Multiple medicated as well as non-medicated approaches have been suggested as antimicrobial and anti-biofilm agents to diminish the activity of the cariogenic oral flora. However, the effectiveness of such agents is still unclear. Also, the cost is an important factor that limits the accessibility of these agents in clinical practice. On the other hand, many salts such as sodium chloride (NaCl), potassium chloride (KCl), and potassium iodide (KI) are readily available and comparatively inexpensive. Hirasawa and Takada [17] reported that adding NaCl to certain antibiotics such as aztreonam and carumonam inhibits S. mutans growth. In addition, it was reported that the combination of sodium salts with certain materials such as sodium bicarbonate (NaHCO3) or human salivary lysosome causes S. mutans cells to lyse [1819]. It was found that when S. mutans was treated with mutanolysin and 1 M NaCl, a significant S. mutans lysis was observed [20]. Moreover, it was demonstrated that S. mutans attachment to salivary agglutinin (which is necessary to initiate the carious lesion) was inhibited by using NaCl or KCl salts [21]. However, some studies indicated that NaCl and KCl were not effective in reducing the cariogenicity or the growth of S. mutans [222324]. A combination of KI and silver diamine fluoride (SDF) was found to diminish S. mutans growth [25]. This combination was able to inhibit caries progression in vitro and prevent biofilm formation [26].
It would be of interest to explore the effect of NaCl, KCl, and KI on the total growth absorbance and biofilm formation of S. mutans and nicotine-induced S. mutans biofilm. Therefore, the aim of this study was to investigate the independent inhibitory effects of NaCl, KCl, and KI salts on nicotine induction of S. mutans biofilm formation.
MATERIALS AND METHODS
Determination of the minimal biofilm inhibitory concentration of the salts
Before beginning the analysis of the effects of the salts on nicotine-induced S. mutans biofilm formation, a preliminary experiment was done to ascertain the minimal biofilm inhibitory concentration (MBIC) of NaCl, KCl, and KI directly on S. mutans biofilm formation without the effect of nicotine. The most commonly used antibacterial concentrations of NaCl, KCl, and KI in the literature range from 0.075 to 0.35 M [171819272829]. Therefore, concentrations of NaCl, KCl, and KI ranging from 0 to 1.8 M were examined. An overnight culture of S. mutans UA159 (ATCC 700610, American Type Culture Collection, Rockville, MD, USA) was grown in tryptic soy broth (TSB) at 37°C in 5% CO2 and stored with 10% glycerol at −80°C for future use.
A 24-hour culture of S. mutans UA159 was grown in TSB and 10 µL was inoculated in microtiter plate wells with 190 µL of 0–1.8 M of NaCl, KCl, or KI in quadruplicate in TSB supplemented with 1% sucrose (TSBS). The microtiter plate was incubated at 37°C in 5% CO2. A spectrophotometer (SpectraMax 190, Molecular Devices Inc., Sunnyvale, CA, USA) was used to determine total bacterial growth by measuring absorbance at 595 nm. The remaining planktonic cells were discarded from the biofilm microtiter plate wells (leaving their attached biofilm), and 200 µL of 10% formaldehyde was added to each well for 30 min to fix and kill the adherent biofilm cells. After 30 minutes, the formaldehyde was removed and the biofilm cells were washed 3 times with deionized water. 200 µL of 0.5% crystal violet dye was added to each well and the biofilm cells stained for 30 minutes. The wells were rinsed 3 times and 200 µL of 2-isopropanol was added to each well for 1 hour to lyse the cells and extract the crystal violet. The plates were read in a spectrophotometer at 490 nm to measure biofilm formation (Figure 1B). Controls included biofilm cells of S. mutans without the NaCl, KCl, and KI salts. The MBIC was established where there was an obvious decrease in the total bacterial growth absorbance. The MBIC of the salts were determined to be 0.45 M for NaCl, 0.23 M for KCl, and 0.113 M for KI.
Inhibition of nicotine-induced biofilm formation by the salts
After determining the MBIC for the three salts, aqueous stock solutions of NaCl, KCl, and KI were prepared for NaCl (4.2 g/40 mL; 1.8 M), for KCl (5.4 g/40 mL; 1.8 M), and for KI (11.95 g/40 mL; 1.8 M). All were stored in a −20°C freezer. An overnight culture of S. mutans UA159 was grown for 24 hours in TSB at 37°C in 5% CO2. NaCl, KCl, and KI salts were diluted in TSBS to obtain the MBIC determined above (0.45 M for NaCl, 0.225 M for KCl, and 0.113 M for KI). The MBIC dilutions of each salt in TSBS were prepared to have 0, 0.25, 0.5, 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 mg/mL nicotine (Sigma-Aldrich Chemical Co., St. Louis, MO, USA). Another identical set of nicotine dilutions in TSBS were prepared without the NaCl, KCl, and KI salts. In addition, each salt was investigated alone. Each nicotine concentration with and without the salt preparation was assayed using the crystal violet biofilm staining procedure described above. Controls involved biofilms of S. mutans without nicotine and with or without NaCl, KCl, and KI salts.
Statistical analysis
Each experiment was performed in triplicate to gain statistical confidence. Each group consisted of 4 wells (n = 4). Two-way analysis of variance (ANOVA) was utilized to compare the effects of NaCl, KCl, and KI salt exposure and nicotine and their interaction on biofilm and total growth absorbance. Pair-wise comparisons were achieved using Fisher's Protected Least Significant Differences method to control the overall significance level at 5%. The primary focus of the study hypotheses was the comparisons involving NaCl, KCl, and KI salt exposure. The distribution of the measurements was investigated and data transformation (natural logarithm, square root, rank, etc.) was used for the analyses to satisfy the ANOVA assumptions. Based on prior studies, the within-group standard deviation of the absorbance measurements for biofilm formation was estimated to be 0.15. With 4 samples in each of 3 replicates of the study, the study power was estimated to be 80% power to detect a difference of 0.2 between salts for each nicotine concentration, assuming 2-sided tests were conducted at an overall 5% significance level.
RESULTS
Determination of minimal biofilm inhibitory concentration of salts on S. mutans
The MBIC of the three salts were determined as following: 0.45 M for NaCl, 0.23 M for KCl, and 0.113 M for KI. Results indicated NaCl, KCl, and KI were able to inhibit biofilm formation significantly (p < 0.05). With NaCl, there was a significant reduction (p < 0.05) in biofilm formation between 0.45 and 0.67 M (Figure 2A). Therefore, 0.45 M of NaCl was recognized as the MBIC for S. mutans biofilm formation in this study. With KCl, most of the concentrations assayed were capable of inhibiting biofilm formation (Figure 2B). However, 0.225 M was selected to be the MBIC for purposes of this study. Finally, 0.113 M of KI was determined as the MBIC for this study as any higher concentration than that was bactericidal (Figure 2C).
 | Figure 2Effect of sodium chloride (NaCl), potassium chloride (KCl), and potassium iodide (KI) on S. mutans biofilm formation. (A) Effect of NaCl on S. mutans biofilm formation; (B) Effect of KCl on S. mutans biofilm formation; (C) Effect of KI on S. mutans biofilm formation.
*Asterisks indicate significant differences compared to samples without the respective salt (p < 0.05).
|
Effect of salts on nicotine-induced biofilm formation
Overall, there was a significant effect for salt presence, nicotine concentration, and their interaction for all salts (NaCl, KCl, and KI) and measures (biofilm and total absorbance). Without salts, nicotine increased biofilm formation especially at 4 and 8 mg/mL. Nicotine demonstrated inhibitory action when the concentration was 16 and 32 mg/mL compared to the group without nicotine (p < 0.05). The NaCl/nicotine, KCl/nicotine, and KI/nicotine interactions demonstrated significant effects inhibiting S. mutans biofilm formation with all concentrations in comparison to the no salt groups except when the nicotine concentration was 32 mg/mL (p < 0.05) (Figures 3, 4, 5). Regarding the total absorbance, minor growth restriction was observed with salts/nicotine interaction. NaCl demonstrated significant inhibition only when the nicotine concentration was 0, 4 and 8 mg/mL (p < 0.05), while KCl demonstrated significant inhibition with 4 and 8 mg/mL of nicotine only (p < 0.05). The greatest amount of total absorbance inhibition was seen with KI. KI inhibited total absorbance growth when the nicotine concentration was 1, 2, 4, 8, and 16 mg/mL (p < 0.05).
 | Figure 3Combined effect of sodium chloride (NaCl) and nicotine on S. mutans total absorbance (A) and biofilm formation (B).
*Asterisks indicate significant differences compared to samples without NaCl (p < 0.05).
|


DISCUSSION
The preliminary experiment was designed to confirm the antimicrobial activities of the three salts and to determine the MBIC of each salt. Even though the antimicrobial activity of these salts was previously reported, there is a lack of evidence regarding the direct and independent effect of these salts on S. mutans biofilm growth. Furthermore, there have been no reports of the effect of these salts on nicotine-induction of S. mutans biofilm. An overnight culture of S. mutans was placed in 96-well microtiter plates with TSBS to facilitate S. mutans growth and biofilm formation. Nicotine was added to both study and control groups to increase biofilm formation as this was confirmed in previous studies [10111213]. The results of the present study indicate that NaCl, KCl, and KI are able to promote reduction in the biofilm formation of S. mutans without nicotine. In addition, these three salts are able to significantly limit the ability of nicotine to increase the growth of S. mutans, especially biofilm formation.
The most important phase of bacterial growth is the biofilm phase as it is the most favorable one for oral bacteria to grow in vivo in order to trigger dental caries demineralization and lesion formation [30]. Also, protein expression in S. mutans biofilm cells differs from that observed in planktonic cells [30]. Therefore, even though the three salts demonstrated minor inhibition of total growth of nicotine-treated cells, this inhibition was recognized clearly when biofilm formation was measured. This may indicate that these salts, in a specific mechanism, decrease the adhesion capabilities of S. mutans. In the nicotine group without salt, 0.25 to 8 mg/mL of nicotine increased S. mutans biofilm formation. The MBIC and MBC were established to be 16 and 32 mg/mL of nicotine, respectively, and this is similar to what was reported by Huang et al. [10]. The rationale is that nicotine is toxic to S. mutans above 8 mg/mL of nicotine. The results of the present study demonstrate that any salt co-incubated with 16 or 32 mg/mL of nicotine demonstrated more inhibition than nicotine alone and this is most likely because of the synergistic antimicrobial effect of the salts and nicotine. But at 8 mg/mL of nicotine, it was observed that with no salt, there was more biofilm formation than concentrations < 8 mg/mL nicotine. However, when any of the salts were added, there was less biofilm formation with 8 mg of nicotine than with concentrations < 8 mg/mL nicotine. A reasonable explanation is that a synergistic effect exists between each salt and nicotine at this specific concentration of nicotine (8 mg/mL).
The potential clinical implication of these results can be related to the amount of nicotine in human saliva of smokers. Feyerabend et al. [8] reported that the nicotine level in human saliva ranges from 0–0.31 mg/mL for those people who are not smokers but are affected by secondary or tertiary smoke. For light or medium smokers, the range of nicotine in human saliva is between 0–1.33 mg/mL and for heavy smokers is 0–2.27 mg/mL. In another study, it was demonstrated that the level of nicotine in human saliva ranges from 0.07–1.56 mg/mL for those people who have smoked for at least 10 years [7]. Supposing the average nicotine level in human saliva of smokers is 1 mg/mL, the present in vitro data indicate that the presence of 0.45 M of NaCl, 0.23 M of KCl, and 0.113 M of KI with 1 mg/mL of nicotine significantly inhibited nicotine-induced S. mutans biofilm formation by 52%, 79.7%, and 64.1%, respectively. However, the actual concentration of nicotine in dental plaque is unknown. Further research may identify how NaCl, KCl, and KI specifically inhibit S. mutans biofilm formation, and what effect these salts have on EPS, Gtfs, GbpA, and lactic acid expression. Nicotine increases EPS, GbpA, Gtfs, and lactic acid production [12]. Therefore, NaCl, KCl, and KI may inhibit EPS, GbpA, Gtfs or lactic acid synthesis. In addition, in vivo studies are needed to confirm the inhibitory effect of these salts and their biocompatibility with oral tissues.
NaCl, KCl, and KI have been used in the medical field for many decades for different purposes such as wound cleaning, nasal irrigation, fungal infections, treatment of hypokalemia, and electrolyte imbalance [313233]. The amount of electrolytes in serum and saliva has been reported in the literature. The normal sodium, potassium and chloride levels in the human serum are 141 mmol/L, 4.3 ± 0.2 mmol/L, and 107.5 ± 2.5 mmol/L, respectively [34]. In saliva, the levels of sodium, potassium, and chlorine are found to be approximately 17.5 mg/dL, 22.6 ± 4.5 mmol/L, and 46.5 mmol/L, respectively [34]. The level of protein-bound iodine in human blood is around 95 µg/L [35]. In the oral cavity, it has become evident that the amount of iodide secreted by salivary glands is higher than in the plasma by 20–100 fold [36]. As this study indicates the possible efficacy of using salts solutions as mouthwashes, it is important to explore their effect on electrolytes balance to avoid causing any health problems. In addition to confirming the antibacterial activities of these salts in vivo, future studies may also discuss the salivary levels of sodium, potassium, chlorine, and iodide before and after the use of these salts solutions.
CONCLUSIONS
This study demonstrates that growth of nicotine-induced S. mutans was diminished in the presence of NaCl, KCl, and KI salts. Because NaCl, KCl, and KI demonstrated inhibitory effects on the growth of S. mutans, this could be beneficial clinically by asking smokers to rinse with aqueous solutions of these salts to diminish the cariogenicity of nicotine-induced S. mutans biofilm formation. In vivo studies are required to confirm this benefit.
 XML Download
XML Download