

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
The 2009 swine origin influenza A H1N1 (pH1N1) became the first influenza pandemic in the 21st century. After the first case of illness reported in Mexico and the and the United States in March and April 2009, this infection spread rapidly and infected humans in over 70 countries within 3 months. The World Health Organization declared the pandemic alert level as phases 5–6 for pH1N1 [1]. The infection rates of pH1N1 differed among age groups, with most of infected patients younger than 25 years old. This may be because of the cross-protection of pre-existing antibodies in the elderly, particularly in those who had experienced other pandemic viruses [12].
A diagnostic kit to test for pH1N1 was developed at the beginning of the outbreak. A rapid diagnostic tests using respiratory specimens for pH1N1 are commercially available. However, negative results in rapid diagnostic tests do not completely rule out infection with pH1N1. Typical serological tests can be used to detect r pH1N1 infection, such as hemagglutination inhibition (HI) assay or microneutralization test (MNT) [34]. Although these traditional methods are sensitive, they are complex and labor-intensive, thus a new active serological surveillance method is required to detect influenza viruses as well as to evaluate antigenic match between circulating viruses and vaccine strains [4].
In the present study, we developed and validated a hemagglutinin 1 (HA1)-specific enzyme-linked immunosorbent assay (ELISA) kit for serological detection of neutralizing antibodies against pH1N1 using both human and mouse serum samples. This ELISA kit can be easily and quickly applied in any basic laboratory. The sensitivity and specificity for detecting HA1 of influenza virus were also demonstrated, as compared to the traditional HI assays and MNT. This HA1-specific ELISA kit could be a useful tool for detecting influenza virus outbreaks and determining vaccine efficacy.
Materials and Methods
Production of recombinant H1N1 HA1 protein (rHA1)
Construction of recombinant plasmid
The HA gene of influenza virus A/New York/3571/2009 (H1N1) was synthesizes based on the sequence in the NCBI Influenza Database, and the synthetic HA (residues 18–344) gene fragment was amplified with adaptor primers having restriction enzyme sequences for BalII (forward) and XbaI (reverse) at the 5′ end of each primer and then cloned into the 6x-His-tagged baculovirus transfer vector pAcGP67A (BD Bioscience, Franklin Lakes, NJ, USA).
Production of rHA1
The recombinant transfer vector pAcGP67A and the BaculoGold were co-transfected into Spodoptera frugiperda (SF9) insect cells according to the manufacturer's instruction (BD Bioscience). A high-titer of recombinant virus was obtained from the cell-culture supernatant after two consecutive subcultures. To obtain recombinant HA1 protein, High-Five cells (2×106 cells/mL) were infected with high-titer of recombinant virus at 27℃, 115 rpm for 3 days then cells were harvested and the recombinant proteins in the supernatant were obtained by ammonium sulfate precipitation and Ni-NTA column purification (#1018, ADAR Biotech, Rehovot, Israel). The protein concentration was measured by bicinchoninic acid assay (Thermo Fisher Scientific, Waltham, MA, USA).
Western blotting
rHA1 was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and the separated protein bands were visualized by Coomassie blue staining and silver staining. The proteins were transferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA, USA) and blocked with 5% skim milk in phosphate buffered saline (PBS) containing tween-20 (PBST, 1× PBS and 0.05% Tween-20) for 1 hour at room temperature (RT). The membrane was incubated with an anti-HA1 mouse monoclonal antibody (1:1,000, ATGen, Seoul, Korea) in 0.2% bovine serum albumin (BSA) in PBST after 3 washes with PBST at RT. The membranes were incubated at 4℃ overnight. Horseradish peroxidase (HRP)-conjugated goat anti-mouse antibody (1:3,000, Bio-Rad) as a secondary antibody in 5% skim milk-PBST was added after PBST washing and incubated for 1 hour at RT. After washing with PBST 3 times, the membranes were developed using a chemiluminiescent substrate (Amersham ECL, GE Healthcare, Buckinghamshire, UK) and exposing the protein side of the membrane to X-ray film.
Human serum samples
A total of 147 subjects was enrolled in this study. Forty-seven patients with influenza infection were confirmed by real-time polymerase chain reaction using nasopharyngeal swabs from patients with presenting fever. Blood samples from the patients were obtained at the time of H1N1 diagnosis. The control group of 30 subjects showed no flu-like symptoms or confirmed H1N1 infection. Samples from 70 subjects were collected before vaccination (pre-vaccination) and resampled at 4 weeks after vaccination with H1N1-inactivated influenza vaccine (post-vaccination). This study was approved by the Institutional Review Board Committee of Gangnam Severance Hospital (Seoul, Korea; IRB No. 3-2009-0170). Informed consents were obtained from all participants and all procedures were carried out in accordance with the relevant guidelines and regulations.
HI assay
The HI assay was performed as described previously [5]. Briefly, sera were treated with receptor destroying enzyme at 30℃ for 20 hours and inactivated at 56℃ for 1 hour. The samples were serially diluted with PBS in the V-bottom 96-well plates then mixed with a standard amount of virus (4 HAU). The plates were incubated for 30 minutes at RT, and 0.5% of chicken red blood cells were loaded into each well and further incubated for 30 minutes at 25℃. The HI endpoint was the highest serum dilution at which agglutination was not observed.
Microneutralization test
Serum neutralizing antibody titers were determined by conducting a multi-well plate cell culture assay. Briefly, MDCK cells were seeded at 1.5×104 cells/well in a 96-well culture plate and cultured at 37℃, and 5% CO2 in an incubator to form a monolayer. Serially diluted samples pretreated with receptor destroying enzyme were mixed separately with 100 TCID50 of A/New York/3571/2009 (H1N1) and incubated at 37℃ for 1 hour. The serum and virus mixture was added to the monolayer of MDCK cells. The plates were further incubated for 18 hours at 37℃ and washed with PBS 3 times, and fixed with 80% acetone for 10 minutes. The presence of viral proteins was confirmed by the monoclonal antibody against influenza nucleoprotein (NP), and the end point of the neutralizing titer was determined as the highest serum dilution at which NP was reduced by 50%.
Enzyme-linked immunosorbent assay
Optimal dilutions of purified H1N1-rHA1 antigen were determined by various conditions and repeated experiments. For antigen coating, the indicated concentration of rHA1 antigens (7.5 ng/mL) was diluted in cold carbonate buffer (pH 9.6) in 96-well microplate and incubated at 4℃, overnight. Antigen-free carbonate the buffer was coated for endpoint assay and the background was subtracted. 3% of BSA in PBST was used for blocking (37℃ for 1 hour). After washing the plate 5 times, serum samples were diluted and pre-incubated with blocking buffer containing supernatant of the High-Five cell culture medium (10 µg/mL of protein concentration) at RT for 10 minutes. Next, 100 µL of diluted serum samples (1:100) was applied in duplicate to each antigen-coated and non-coated well and incubated for 90 minutes at 37℃. After the plates were washed 5 times with PBST, HRP-conjugated rabbit anti-human IgG (1:10,000, Sigma-Aldrich, St. Louis, MO, USA) and anti-mouse IgG Ab (1:5,000, Santa Cruz Biotechnology, Dallas, TX, USA) were added to each well matching the sample origin. Next, the plates were incubated for 90 minutes at 37℃ and washed 7 times with PBST. For color development, 100 µL of 3,3′,5,5′-tetra-methylbezidine in acidic buffer (KPL, Gaithersburg, MD, USA) was added to each well and incubated for 15 minutes at RT. The reaction was stopped by adding an equal volume of 2.5 N H2SO4. The optical density (OD) was determined at a wavelength of 450 nm using a microplate reader within 30 minutes. Endpoint titers of human H1N1-HA1-specific Abs were expressed as the OD450 value of rHA1 uncoated well subtraction from rHA1-coated wells.
Immunogenicity and protective efficacy in a murine model
Animals
Six-week-old female C57BL/6 mice were purchased from Orient Bio (Seoul, Korea). The animals were maintained in the animal care facilities of the Korea Research Institute of Bioscience and Biotechnology (KRIBB) under specific pathogen-free conditions. The experiments were approved by the Institutional Animal Care and Use Committee of KRIBB (KRIBB-AEC-14058).
Vaccination, sample collection, and viral challenge
Each group of mice was immunized with 0.2, 0.5, or 0.8 µg of influenza virus A/California/04/2009 (CA04) vaccine candidate, suspended in a volume of 0.2 mL by intramuscular injection twice at a 2-week interval. For the control experiment, 0.2 mL of PBS was injected into the mice intramuscularly. After 2 weeks after the last vaccination, the mice were anesthetized with a Zoletil 50 mix (0.015 mL/20 g, Virbac Laboratory, Carros, France) and Rompun injection (0.5 mg/20 g, Bayer Korea, Seoul, Korea). Sera were collected by orbital bleeding. Next, the mice were challenged with 10× the LD50 of live influenza A virus in 30 µL of PBS into intranasal route. The body weight change and survival rate of the animals were also evaluated.
Statistical analysis
All data were expressed as the mean±standard deviation. Statistical comparisons between experimental groups were performed using the Student's t test. The sensitivity and specificity of ELISA for detecting the anti-HA1 specific antibody were reported by calculating the area under the receiving operator characteristic curve. The correlation between the anti-HA1 antibody and HI titer was determined by Pearson correlation test. GraphPad Prism 8 (GraphPad Software Inc., La Jolla, CA, USA) was used for all analyses. p<0.05 was considered statistically significant.
Results
Recombinant HA1 proteins were expressed in the baculovirus insect cell system
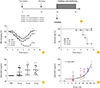
Recombinant H1N1-HA1-18-344 (rHA1) was expressed in the baculovirus insect system (Fig. 1A, B). Proteins were produced by High-Five cells after vector transformation. Purified rHA1 protein was confirmed by Coomassie blue and silver staining (Fig. 1C). H1N1 proteins of the expected size of 46 kDa were isolated.
The ELISA system using recombinant HA1 protein was established to detect anti-rHA1 antibody in patients with influenza virus infection
The ELISA system using rHA1 protein was developed and validated to detect anti-rHA1 antibodies in the serum samples from influenza-infected patients and controls. According to this ELISA, OD values of patients were increased than controls (0.942±0.371 vs. 0.212±0.086, p<0.001) (Fig. 2A). And in vaccinated individuals, anti-rHA1 antibody levels were significantly increased after vaccination compared with levels from pre-vaccination status (Fig. 2B). From these data, this ELISA system using rHA1 could be useful in diagnosis of influenza infection and in discrimination of vaccination status in human.
The ELISA system was compared to standard detection methods for influenza virus infection, such as the HI assay and MNT. As shown in Fig. 2C, this ELISA system could be a sensitive and specific method for detecting the anti-rHA1 antibody in influenza-infected patients. The detection performance of this ELISA system was comparable to that of the HI assay and MNT.
Anti-rHA1 antibodies were detected by the ELISA system in the animal model of influenza virus infection
The ELISA system using rHA1 protein was also validated in an animal model. The control (PBS) and vaccinated groups were compared. The vaccinated groups were immunized with influenza virus A/California/04/2009 (CA04) vaccine. The vaccines contained 3 different amounts of antigens, such as 0.2, 0.5, and 0.8 µg in each single dose of vaccine, respectively. All mice were immunized with PBS or vaccines twice in 2 weeks apart and challenged at 2 weeks after the last vaccination (Fig. 3A). Body weight changes and the survival rate of vaccinated and non-vaccinated groups were monitored after influenza virus challenge (Fig. 3B, C). Compared to the control PBS group, the vaccinated groups showed lower body weight changes (Fig. 3B) and death rates (Fig. 3C). The serum levels of anti-rHA1 antibodies were measured in both non-immunized and immunized mice by ELISA. The increased levels of antibodies were detected in all vaccinated mice but not in PBS group. The amount of antigens in vaccines did not affect the production of anti-rHA1 antibodies in this study (Fig. 3D). And the levels of anti-rHA1 antibodies were significantly correlated with the results of HI assay using the same samples (r=0.508, p<0.001) (Fig. 3E).
Taken together, anti-rHA1 antibodies were detected by the ELISA system in vaccinated mice. The positive correlation between anti-rHA1 antibody measured by ELISA and titers in HI assay was also observed in the animal model.
Discussion
A striking influenza pandemic occurred in 2009–2010. This highly pathogenic virus came from swine and the next pandemic virus has been thought to have arisen from avian sources in Asia [4]. Because of unpredictable and diverse sources, techniques for global surveillance in influenza virus infection that could be used both in humans and animals has been required. Humans have been exposed to influenza viruses that underwent antigenic and genetic changes; thus, assays that are strain-specific have been also required to characterize the viruses and detect immune responses to recent infection or vaccination [4]. The standard HI assay or MNT for measuring antibody responses to influenza virus surface antigens were developed previously [67]. Therefore, we focused on being strain-specific, convenient and readily available test for both humans and animals.
In this study, we produced recombinant HA1 influenza protein (2009 H1N1 pandemic strain) using a baculovirus-insect cell system. We established the ELISA system using rHA1 for detecting an anti-rHA1-specific antibody in humans, as well as in mice. Its clinical performance was also demonstrated by the comparison with those of HI assay and MNT. This ELISA system could be used for evaluating influenza virus vaccine efficacy, not even for diagnosing current infections.
Diagnostic techniques for early and accurate detection of newly emerging viruses are important for the rapid initiation of therapy and prophylaxis to control infection during disease outbreaks [6]. Traditional methods for detecting the neutralizing antibodies induced by influenza virus infection are time-consuming and the results could be affected by several external factors [48]. Rapid diagnostic kits for influenza viruses are commonly used to detect influenza infection. The results of rapid diagnostic kits could be completed within 30 minutes and are easily interpreted. However, rapid diagnostic kits showed lower sensitivity (62.3% reported in a meta-analysis) compared reverse transcription polymerase chain reaction-based approaches in the 2009 pandemic season. Accurate diagnosis of new emerging pathogen was limited at that time [6].
In contrast, the ELISA system developed in this study, could be used to detect specific antibodies against certain strain of influenza virus in a short time. This method is relatively easy and convenient compared to the HI assay or MNT, with similar high sensitivity and specificity for diagnosis of influenza virus infection compared to the HI assay or MNT. Our method does not require a live virus to perform or complex procedures. Moreover, this ELISA system could discriminate the vaccination status of the subjects, and thus the efficacy of vaccines could be also evaluated.
In summary, we established a recombinant HA1-based ELISA system for detecting anti-rHA1 antibody against influenza virus infection in humans. We developed a diagnostic approach for influenza virus infection which showed good clinical performance. This method is also applicable for evaluating the vaccination status of the patients and efficacy of influenza virus vaccines. The ELISA system developed in this study is a useful approach for diagnosis of influenza virus infection and assessment in vaccine efficacy.


 XML Download
XML Download