

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Creation of an embryo via the somatic cell nuclear transfer (SCNT) method has various potential applications such as species conservation [21], regenerative medicine [10], and animal husbandry [38]. After the initial production of the cloned sheep ‘Dolly’ by Campbell et al. [7], cloned animals were developed by applying SCNT in 23 mammalian species including mouse [37], bovine [15], pig [3], and primate [68]. However, the use of SCNT has several problems. Above all, low efficiency is the biggest obstacle in the application of SCNT [17]. Many previous studies have shown success in inducing developmental rate improvement by using chemicals [1335], vitamins [22], and histone modifiers [511]. Although some groups have elevated the developmental rate to near 90% by using histone deacetylase mRNA, an accurate description of the developmental pathways in SCNT murine embryos is unreported [23].
Polo-like kinase 1 (Plk1) is also known as serine/threonine-protein kinase 13 (STPK13) [12]. It is encoded by the PLK1 gene in mouse chromosome 7 and human chromosome 16. Plk1 is considered a proto-oncogene because it is observed frequently in tumor cells [20]. A defect in Plk1 expression causes hindrance of cell growth via a pro-apoptotic pathway [4]. In somatic cells, Plk1 localizes with centrosomes in the interphase [18]. Plk1 regulates cytokinesis including centrosome and chromosome maturation [36], adhesion of kinetochore-microtubules, and timing of mitotic division initiation [24]. During meiosis, Plk1 acts as an essential microtubule-associated protein together with anaphase-promoting complex and CDC5 [31].
Many studies have demonstrated that Plk1 has a critical role in mitotic cell division in somatic cells and cancer cells [92930]. Most recently, some studies have shown the importance of Plk1 in mammalian meiotic and early mitotic stage [162739]. Spindle organization proteins, such as Plk1, also have critical roles in meiotic and mitotic divisions including chromosome segregation in the pre-implantation stage of mouse embryos. Compared with in vivo-fertilized embryos during early cleavage, abnormal chromosome segregation is reported to occur frequently in SCNT murine embryos [25]. Thus, the expression and localization patterns of Plk1 in SCNT murine embryos were investigated and compared with in vivo counterparts to demonstrate the correlation between Plk1 expression and poor development of SCNT murine embryos. Although, some studies have demonstrated that Plk1 is an essential factor in mammalian embryos, in SCNT murine embryos, the expression patterns of Plk1 have not been reported. Therefore, this study investigated the Plk1 expression patterns in SCNT murine embryos.
Materials and Methods
Animal use and ethical statement
All metaphase II (MII) oocytes and donor cells were acquired from 6 to 8 weeks old C57BL6 X DBA2 F1-hybrid (B6D2F1) female mice (Orient Bio, Korea). Animal experiments were approved under the agreement guidelines of the Institutional Animal Care and Use Committee of Seoul National University (approval No. SNU-130123-5-5).
Collection oocytes and preparation of donor cells
The 7.5 IU of equine chorionic gonadotropin (eCG; Daesung Microbiology Labs, Korea) were introduced to female B6D2F1 mice by intraperitoneal injection for superovulation. Forty-eight hours later, 7.5 IU of human chorionic gonadotropin (hCG; Daesung Microbiology Labs) were injected into the mice. To obtain in vivo-fertilized embryos, two female mice were mated with a male mouse immediately after hCG injection. Oocytes and in vivo-fertilized embryos were collected 15 h after the hCG injection. Briefly, oviducts of mice were transferred to 2 mL TCM-199 with Earl's salts medium (TCM-washing) including 300 IU/mL of hyaluronidase to remove cumulus cells. Oocytes were recovered from torn ampullae. After 3 to 4 min of exposure to the hyaluronidase-containing TCM-washing medium, the cumulus-free oocytes were washed five times in Hepes-CZB medium (HCZB) before manipulation. The cumulus cells were suspended in a 10 µL TCM-washing droplet mixed with 12% polyvinylpyrrolidone (PVP) for subsequent use as donor cells. Finally, the entire dish was covered with mineral oil.
Enucleation
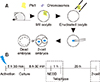
Oocytes (12–15) were set in a 10 µL droplet of HCZB, including 10 µ and kept in a 37℃ operation heat chamber on a microscope stage. The zona pellucida of the oocyte was pierced by an enucleation pipette with several piezo pulses using a piezo-actuated micromanipulator (PMM-150FU; Prime Tech, Japan). The diameter of the enucleation pipette was 6 to 8 µm. The MII chromosome-spindle complex was then removed via enucleation pipette (panel A in Fig. 1). The enucleated oocytes were transferred to HCZB, washed three times, and kept for up to 30 min in KSOM medium.
Nuclear injection
The cumulus cells were collected by using 5 µm diameter injection pipettes. The cells were aspirated in and out several times to accomplish membrane destruction. The zona pellucida was drilled out by an injection pipette and piezo pulses. Then, the donor cells in the pipette were injected into the enucleated oocytes by a single piezo pulse. After that SCNT, all of the reconstructed oocytes were washed three times in HCZB and kept for 10 min in room temperature HCZB to stabilize the oocytes. All injection protocols were undertaken at room temperature. After oocyte stabilization, reconstructed oocytes were kept in KSOM in a humidified 37℃ incubator.
Activation and culture
The embryo culture medium was based on KSOM and CZB. Reconstructed SCNT oocytes and MII oocytes were kept separately in calcium-free CZB with 5 µg/mL cytochalasin B (CB) and 10 mM SrCl2 (ACZB) for oocyte activation. After 5 h 30 min, activated oocytes were washed in HCZB five times and transferred in KSOM medium to a humidified 37℃ incubator. The in vivo-fertilized embryos were similarly cultured in KSOM medium until analysis.
Treatment of a Plk1 inhibitor, BI2536
As Plk1 is essential for mitotic division in in vivo-derived embryos, embryos were treated with 2, 4, 10, 20, 100, or 500 nM of BI2536 to investigate the optimal concentration required for Plk1 inhibition. The embryos used in the control group were cultured in KSOM medium with 1% dimethyl sulfoxide. The embryos used in the experimental groups were placed in KSOM medium with the indicated concentrations of BI2536. All embryos were cultured in a 37℃ humidified incubator for 30 min. Immunofluorescence analysis was performed at the times in the boxes with dotted lines as described in panel B in Fig. 1.
Immunofluorescence, live imaging and image analysis
The reconstructed oocytes were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 30 min, then the oocytes were washed three times in PBS containing 0.5% PVP and 0.1% Triton X-100 washing buffer. The oocytes were placed in PBS containing 0.25% Triton X-100 for 4 h to make the membrane permeable and, then, incubated in PBS with 0.1% Triton X-100 and 1% BSA for blocking for 2 h in a 37℃ incubator. Plk1 was detected by rabbit polyclonal IgG antibodies (1:100; Santa Cruz Biotechnology, USA) and goat-anti-rabbit polyclonal IgG antibodies (1:500; Millipore, USA). DAPI staining was used to assess nuclear morphology. Confocal microscopy images were acquired by using an LSM 700 microscope (Zeiss, Germany). A JuLI Stage recorder (NanoEntek, Korea) was used to obtain live cell images. The confocal microscopy images were analyzed using Zen2 BLUE edition software (Zeiss).
Statistical analysis
All experiments were iterated three times at least. All percentage data obtained in this study are presented as mean ± SD values. The embryonic development and first mitotic division data were analyzed by applying Student's t-test. A probability of p < 0.05 was considered significant.
Results
Comparison of first mitotic division efficiencies of SCNT murine embryos and in vivo-fertilized embryos
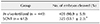
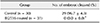
The success rate for the first mitotic division of SCNT murine embryos (53.1 ± 2.3%) was significantly lower than that of in vivo-fertilized embryos (90.9 ± 5.9%; p < 0.05) (Table 1). The spindle and chromatin together with adjacent cytoplasm were removed by an enucleation pipette during the SCNT process, resulting in the removal of the greatest proportion of spindle-binding proteins including Plk1. The loss of Plk1 may have caused the low mitotic division rate of the SCNT murine embryos. Consequently, experiments were designed to analyze the expressions of Plk1 before and after mitosis in both SCNT and in vivo-fertilized murine embryos.
Intensity of Plk1 was significantly lower in enucleated oocytes than in MII oocytes
The fluorescence intensity of Plk1 expression was measured by performing immunofluorescence analysis. In MII oocytes, marked fluorescence intensity of Plk1 was observed around the chromosomes and the spindle apparatus (panel A in Fig. 2; green). However, enucleated oocytes had low Plk1 fluorescence intensity as Plk1 was removed with the chromosomes during the enucleation process (panel B in Fig. 2). Quantization data obtained by confocal microscope analysis showed that the fluorescence intensity of Plk1 in MII oocytes was over five times higher than the intensity of Plk1 in enucleated oocytes (panel C in Fig. 2).
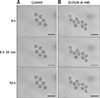
Mitotic division of embryos was blocked by BI2536, a Plk1 inhibitor
The images in panels A–G in Fig. 3 show the morphology of embryos that were treated with different concentrations of BI2536. About 60% of the 2 nM BI2536-treated embryos and more than 90% of the untreated in vivo-fertilized embryos developed into 2-cell embryos, whereas all embryos treated with more than 4 nM of BI2536 failed to attain 2-cell division (panel H in Fig. 3). Further, the groups of embryos that were treated with highs concentrations of BI2536, such as 100 nM and 500 nM, were dead within 30 min. The dead embryos showed membrane collapse and shrinkage (panels F and G in Fig. 3). Based on these results, 4 nM of BI2536 was applied to block Plk1 in the next series of experiments. In addition, the effect of BI2536 was tested in parthenogenetic murine embryos. All parthenogenetic embryos that were treated with 4 nM of BI2536 did not develop to 2-cell embryos (0.0 ± 0.0%), while most untreated embryos developed to the 2-cell stage (96.7 ± 4.3%) (Table 2, Fig. 4). Taken together, the results above show that BI2536, a Plk1 inhibitor, causes blocking of mitotic division in both in vivo-fertilized and parthenogenetic murine embryos. The experimental results confirm that Plk1 has a critical role in mitosis in early-stage mouse embryos. Therefore, in the following experiments, the focus was placed on the localization of Plk1 within the mouse embryo.
Abnormal expression pattern of Plk1 was shown in SCNT murine embryos with developmental failure
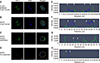
From fertilization to the 2-cell stage, the dual immunofluorescence labeling images showed that Plk1 was located around the nuclei in embryos that developed normally (panel A in Fig. 5). These results show that Plk1 gathers around the nuclear membrane from fertilization to the 2-cell stage under normal conditions. In addition, Plk1 expression was present on the nuclear membrane in 2-cell stage embryos. Interestingly, Plk1 exhibited a bridge-like morphology by being present between the two nuclei in 2-cell stage embryos with normal development. However, the SCNT murine embryos, which failed to reach the 2-cell developmental stage, presented two notable Plk1 results: ectopic Plk1 localization and low Plk1 expression. Among the in vivo embryos, 94% showed normal Plk1 expression patterns with only 6% of those embryos showing a low Plk1 expression pattern. However, among the SCNT murine embryos, the low Plk1 expression pattern was twice that in the in vivo-fertilized group. In addition, the ectopic pattern, in which nuclei and Plk1 proteins were not co-located, was observed in the 35.2% of the SCNT murine embryos (panel B in Fig. 5, Table 3).
Next, to describe Plk1 expression in more detail, Plk1 and DNA were double-stained and monitored by using confocal microscopy in both in vivo-fertilized and SCNT murine embryos. Images of 2-cell stage in vivo-fertilized embryos showed a bridge-like Plk1 expression morphology by Plk1 expression (panel A in Fig. 6). The Plk1 intensity levels in embryos with developmental failure were less than 1.0e04, while those in normally developed embryos were 2.0e04 and above, which is twice higher, than in developmental failure group. Based on those results, embryos with a Plk1 intensity value under 1.0e04 were classified as low intensity. The SCNT 2-cell embryos that developed normally also exhibited a bridge-like Plk1 expression morphology (panel B in Fig. 6). However, compared to the normally developed embryos, the SCNT murine embryos that failed to develop to the 2-cell stage exhibited the two Plk1 expression pattern types: ectopic Plk1 expression (panel C in Fig. 6) and low Plk1 expression (panel D in Fig. 6). The Plk1 expression intensity was higher in the nuclear membrane boundary region than on the other side of the cytoplasm (panels E and F in Fig. 6; arrowheads). In the group showing Plk1 ectopic expression, the expression of Plk1 was significantly lower in the nuclear membrane boundary (arrowheads) than on the other side of the cytoplasm (panel G in Fig. 6). Twelve percent of the SCNT murine embryos had a low-intensity Plk1 expression pattern (panel H in Fig. 6). Taken together, the Plk1 expression location in the embryos that developed to the 2-cell stage normally, including SCNT embryos, was around the nuclear membrane boundary in both 1-cell and 2-cell stage embryos. However, the Plk1 expression of the SCNT murine embryos that failed to develop showed both a low level of Plk1 expression and ectopic Plk1 expression.
Discussion
In previous studies, Plk1 has been shown to be an essential element in mitotic cell division [14]. However, cell division of embryos during development is different from that of somatic cells [26]. Recently, some studies have demonstrated that Plk1 is a critical protein in meiosis and at the embryo pre-implantation stage [240]. Mouse and primate SCNT procedures include an enucleation process that removes the MII spindle. This method results in the loss of chromosomes and spindle-binding proteins. However, oocytes and embryos before the 2-cell stage do not have an intact zygotic genome activation ability [34]. Accordingly, protein rescue of early-stage embryos depends on maternal mRNA. Therefore, early-stage SCNT murine embryos have an inadequate protein rescue ability for chromosomes and spindle-binding proteins. Hence, we focused on the effect of Plk1 loss during the development of SCNT murine embryos in this study.
First, our results confirmed that Plk1 was located together with the chromosomes in MII stage oocytes. Consequently, after enucleation, there was a low Plk1 expression in the enucleated oocytes. This result shows that Plk1 is removed with the chromosomes during the enucleation process. Next, the embryos were treated with BI2536, a Plk1 inhibitor, to determine whether BI2536 affects early mitotic division in mouse embryos. Although the 50% inhibitory concentration value of BI2536 is reported to be 0.83 nM in human cancer cells [32], some research groups have used higher concentrations of BI2536 such as 100 to 500 nM to inhibit mitosis [119]. In this study, BI2536 treatments over 100 nM produced rapid apoptosis of embryos similar to results reported by another research group studying the toxicity of BI2536 [28]. In this study, 4 nM of BI2536 was sufficient to block in vivo-fertilized and parthenogenetic embryos from reaching the 2-cell stage. Taken together, these results confirm that Plk1 is a crucial cytokinesis protein in mouse pre-implantation embryos.
For verification of the location and expression of Plk1, assessment of immunofluorescence assay results for Plk1 and DNA followed. It was observed that Plk1 was expressed around the nuclei in all of the normally developed embryos. However, the embryos that failed to reach the 2-cell stage exhibited two difference in Plk1 expression patterns; first, there was a low level of Plk1 expression in the embryo's cytoplasm and second, there was a pattern of ectopic Plk1 expression. In those embryos, the Plk1 was not co-located with the nucleus. Ectopic expression of Plk1 was only observed in the SCNT murine embryos that did not develop past the 2-cell stage. These results suggest that ectopic localization of Plk1 might be the result of removing Plk1 recruiting elements, such as Aurora family kinase, inner centromere protein, and survivin [33].
In conclusion, even though the accurate mechanism involved in SCNT murine embryo development remains incompletely described, the results of this study demonstrate that Plk1 has a critical role in the first mitotic division in mouse SCNT murine embryos. In particular, this study suggests the nuclear membrane boundary localization of Plk1 is crucial for successful development in SCNT murine embryos. These results provide a new insight to elucidating the molecular mechanisms in SCNT murine embryos.




 XML Download
XML Download