

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a chronic inflammatory disease of the lung characterized by airway hyperresponsiveness (AHR), chronic airway inflammation, a preferential Th2-immune profile and increased allergen-specific immunoglobulin (Ig) E production. Although the prognosis of asthma is generally good, it caused approximately 397,100 deaths in 2015 globally.1 One of the most useful therapeutic approaches for airway allergic disease is an immunomodulatory intervention aiming to inhibit Th2 immune responses.2345 The hygiene hypothesis states that a reduction in microorganism load in the environment may increase the chance of developing asthma.67 In this regard, for the treatment of asthma and other allergic diseases, immunomodulatory bacterial components may exert potential therapeutic effects by modifying ongoing disease processes or preventing the progression of allergic disease.
Flagellin is the main component of the bacterial flagellum. Flagellin monomers activate the Toll-like receptor 5 (TLR5) signaling pathway, resulting in diverse biological effects. Flagellin B (FlaB) is the main building block of the Vibrio vulnificus flagellar shaft and has a high affinity for TLR5.8 FlaB ameliorates experimental asthma in ovalbumin (OVA)- and house dust mite (HDM)-induced mouse models.91011 Treatment with a high dose of FlaB mixed with OVA mitigates experimental asthma by inhibiting Th1/Th2/Th17 responses in a TLR5-dependent manner by generating regulatory dendritic cells (rDCs) and regulatory T cells (Tregs).9 FlaB treatment also generates rDCs in peripheral blood mononuclear cells (PBMCs) from HDM-sensitive patients with asthma, subsequently suppressing HDM-stimulated Th1/Th2 responses, and enhances Tregs in an interleukin (IL)-10-dependent manner.9 Moreover, FlaB modulates the function of invariant natural killer T (iNKT) cells in response to IL-10 produced by dendritic cells (DCs) in PBMCs derived from patients with asthma.10 These findings suggest that FlaB can be used as an effective immunomodulatory adjuvant for the treatment of allergic asthma.
HDM is globally ubiquitous in human habitats and is a prevalent factor underlying allergic rhinitis and allergic asthma, making it one of the most important sources of indoor allergens.121314 Our previous studies have shown that a HDM/FlaB mixture ameliorates HDM-induced asthma. We wanted to know whether the conjugation of HDM with FlaB or their fusion proteins would have a therapeutic effect on allergic asthma, since FlaB-antigen fusion proteins have shown higher efficacies than mixtures in our previous vaccine studies.1516 Given that HDM preparations comprise a mixture of multiple allergens, it was difficult to standardize a protocol for the conjugation of HDM allergens with FlaB or to create a uniform fusion protein formulation. The HDM allergen Derp2 is the major inducer of allergy and atopic asthma worldwide.17181920 Specific IgE antibodies to Derp2 have been found in 79.2% of patients who have asthma, wheezing and/or rhinitis.2122 Derp2 is a strong risk factor for asthma in children and adults.23 Therefore, the Derp2 protein should be a feasible vaccine candidate for asthma immunotherapy. Immunotherapy using a DNA vaccine encoding the derp2 allergen has shown a suppressive effect in mouse models of Derp2-induced airway inflammation.2425 In the present study, we constructed a Derp2 recombinant protein and Derp2 fusion proteins with FlaB and tested whether the fusion protein vaccines were more efficacious anti-asthma immunotherapeutics. We compared the therapeutic effects of the Derp2-FlaB fusion protein with those of a mixture of Derp2 and FlaB in a Derp2-induced asthma model and found that the Derp2-FlaB fusion protein more effectively suppressed allergic airway inflammation than the Derp2 + FlaB mixture.
MATERIALS AND METHODS
Mice
Eight-week-old female BALB/c mice were bred in a specific pathogen-free facility and housed in autoclaved cages with autoclaved bedding. The Committee on Animal Welfare at Chonnam National University Medical School approved the study protocol.
Expression vector construction and recombinant protein purification
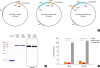
The DNA sequence encoding derp2 was synthesized by the Bioneer Company (Daejeon, Korea). Codon usage was optimized for the expression in Escherichia coli. The synthesized derp2 DNA fragment was cloned into the plasmid pET30a(+) (Novagen, Madison, WI, USA) between the NdeI and XhoI sites for expression as a His-tagged protein (Fig. 1A). The DNA fragment encoding the entire open reading frame (ORF) of flaB from V. vulnificus CMCP6 was fused to the C-terminus or N-terminus of the derp2 DNA fragment and expressed as a Derp2-FlaB or FlaB-Derp2 fusion protein, respectively (Fig. 1A). DNA sequences in the constructs were confirmed by the dideoxy-chain termination method (Macrogen, Seoul, Korea). The resulting expression plasmids were transformed into competent E. coli BL21. Protein expression was induced with 0.2 mM isopropyl-β-D-thiogalactoside (IPTG) overnight, and cells were pelleted by centrifugation and stored at −80°C until use. Cells were lysed by solubilizing the pellets in 6 M guanidine hydrochloride, 100 mM NaH2PO4, 10 mM Tris base and 10 mM imidazole at pH 8. After centrifugation at 18,000 rpm for 30 minutes, cell-free supernatant was loaded on a column containing Ni-NTA agarose beads (Qiagen, Hilden, Germany). The column was washed with washing buffer (100 mM NaH2PO4, 150 mM NaCl, 8 M urea, 20 mM imidazole, pH 8), and proteins were eluted with washing buffer supplemented with 230 mM imidazole. Refolding of the Derp2 and fusion proteins was accomplished by dialyzing the eluted proteins with distilled water. The purity of the recombinant proteins was confirmed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot analysis with an anti-FlaB antibody raised in rabbit. Lipopolysaccharide (LPS) contamination was removed by treatment with TritonX-114 (Sigma-Aldrich, St. Louis, MO, USA),26 and the residual LPS content was determined by using a gel-clotting Endosafe LAL kit (Charles River Endosafe, Charleston, SC, USA). The LPS levels in protein preparations were kept below the Food and Drug Administration (FDA) guideline (less than 0.15 EU/30 g per mouse).
Fig. 1
Recombinant protein construction and analysis. (A) Map of expression vectors used to manufacture the Derp2, Derp2-FlaB and FlaB-Derp2 recombinant proteins. DNA fragments of derp2, flaB, the T7 promoter region, kanamycin resistance gene (Km), and cloning restriction enzyme sites are shown. (B) SDS-PAGE and Western blot analysis of the bacterially expressed and purified Derp2, Derp2-FlaB and FlaB-Derp2 recombinant proteins. The Derp2-FlaB and FlaB-Derp2 proteins were probed with a FlaB antibody. (C) TLR5-mediated NF-κB luciferase activity assay with FlaB, Derp2-FlaB, or FlaB-Derp2 fusion protein. Analysis was carried out using a dual-luciferase reporter assay system, and results were normalized to pCMV-β-galactosidase as a control. All values represent the mean plus the standard error of at least 3 independent experiments.
FlaB, flagellin B; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; TLR5, Toll-like receptor 5; FlaB, flagellin B.
*P < 0.001 compared with FlaB-only.
NF-κB luciferase reporter assay
TLR5-stimulated NF-κB activity was estimated as previously described.15 Briefly, HEK293T cells were transfected with p3XFlag-hTLR527 and pNF-κB-Luc plasmids28 using the Effectene kit (Qiagen). Luciferase activity was normalized to LacZ expression using the control expression plasmid pCMV-β-Gal (BD Biosciences Clontech, Palo Alto, CA, USA). Twenty-four hours after transfection, the cells were incubated with proteins at the same molar ratio for 24 hours. Cells were lysed with a Lysis Buffer (Promega, Seoul, Korea), and luciferase activity was measured by a luminometer (MicroLumatPlus LB 96V; Berthold, Wilbad, Germany).
Asthma model and treatment schedule
The mice were sensitized by intraperitoneal injection of 10 μg of Derp2 and 10 μg of HDM extract adsorbed onto 2 mg of alum on days 0, 7 and 14 as previously reported.25 The mice were then challenged intranasally with 15 μg of Derp2 in 40 μL of phosphate-buffered saline (PBS) for 7 days, from days 25 to 31 under anesthesia. The normal control group received only PBS. To determine the anti-asthmatic adjuvant effect of FlaB, mice were intranasally administered 4 μg of Derp2, a mixture of 4 μg of Derp2 with 12 μg of FlaB, 16 μg of Derp2-FlaB fusion protein or 12 μg of FlaB on days 4, 11 and 18. On day 32, noninvasive AHR was determined, and on day 33, BALF, bronchial LN cells, spleen, serum and lung tissues were obtained (Fig. 2).
Fig. 2
Experimental protocols. BALB/c mice were sensitized by intraperitoneal injections of Derp2 + HDM in alum on days 0, 7 and 14. On days 4, 11 and 18, the mice were treated intranasally with Derp2, Derp2 + FlaB, Derp2-FlaB fusion protein or FlaB under anesthesia. The mice were challenged daily by the intranasal administration of Derp2 on days 25 to 31. The control animals were treated and challenged with PBS. Parameters of allergic responses were evaluated 24 hours after the final Derp2 challenge.
HDM, house dust mite; FlaB, flagellin B; PBS, phosphate-buffered saline; AHR, airway hyperresponsiveness; BAL, bronchoalveolar lavage.

Measurement of noninvasive AHR
Noninvasive AHR was measured using an OCP 3000 barometric plethysmographic chamber (Allmedicus, Anyang, Korea), as previously described.29 Briefly, we prepared methacholine (Sigma-Aldrich) at the following concentrations: 6.25, 12.5, 25 and 50 mg/mL. Mice inhaled each concentration of methacholine for 3 minutes as an aerosol prepared with a NE-U17 (OMRON, Milton Keynes, UK), and enhanced pause (Penh) values were measured for 5 minutes. Airway resistance was expressed in Penh. Data are expressed as the percentage increase in Penh after challenge with each concentration of methacholine, where the baseline Penh (after PBS challenge) was set at 100%.
Collection and analysis of bronchoalveolar lavage (BAL) fluid
To investigate cellular infiltration into the airways, the trachea was exposed and cannulated, and the lungs were lavaged twice with 0.5 mL of PBS. The total volume of BAL fluid was recorded. BAL fluid was centrifuged at 1,500 rpm for 5 minutes at 4°C, and the supernatant was stored at −80°C. The cell pellet was resuspended in 0.5 mL of PBS, the total cell number was determined, and the cells were spun onto glass slides by cytocentrifugation and stained with Diff-Quik solution (Sysmex, Kobe, Japan). The relative numbers of different types of leukocytes were determined from the slides. Data are expressed as the number of cells per mL of BAL fluid.
Cytokine measurement
The cytokines IL-4, IL-5 and IL-13 were measured in BAL fluid using mouse IL-4, IL-5 and IL-13 Ready-SET-Go! ELISA kits (eBioscience, San Diego, CA, USA) in accordance with the manufacturer's protocols.
Measurement of Derp2-specific serum IgE
Derp2 was coated on 96-well ELISA plates (Corning Incorporated, Corning, NY, USA) at a concentration of 1 µg/mL. After blocking, wells were incubated with mouse sera at a 1:30 dilution for 2 hours at room temperature. For detection, anti-mouse IgE-specific horseradish peroxidase (HRP)-conjugated secondary antibodies (Southern Biotech, Birmingham, AL, USA) at a dilution of 1:2,000 were added to the wells. Antigen-specific antibody concentrations were calculated based on a mouse IgE standard (BD Pharmingen; catalog No. 554118).
Histology
Whole lungs were flushed with PBS injected into the right ventricle, removed and fixed in 10% formaldehyde, and then embedded in paraffin. Hematoxylin and eosin (H&E) staining was performed for general morphology, and slides were examined by optical microscopy.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 5.0 software (GraphPad Software, Inc., San Diego, CA, USA). Comparisons were performed with the Mann-Whitney U test. A P value of < 0.05 was considered statistically significant. All data are expressed as the means ± standard error of mean.
RESULTS
Protein purification and analysis
Derp2, Derp2-FlaB and FlaB-Derp2 were expressed as recombinant proteins in E. coli BL21 and purified under denaturing conditions using Ni-NTA agarose beads. Derp2 recombinant protein showed a 14-kDa band by SDS-PAGE, while the Derp2-FlaB and FlaB-Derp2 fusion proteins showed 56-kDa bands by SDS-PAGE, which were also detected by Western blot analysis using a FlaB antibody (Fig. 1B). Luciferase reporter assays were performed to test the TLR5-stimulating activity of the Derp2-FlaB or FlaB-Derp2 fusion protein using 293T cells transiently co-transfected with plasmids harboring the human TLR5 cDNA, an NF-κB luciferase reporter, and beta-galactosidase. The Derp2-FlaB fusion protein significantly enhanced the activation of NF-κB through TLR5 compared with that induced by FlaB alone (11-fold increase) at equimolar concentrations, while the FlaB-Derp2 fusion protein induced significantly less (P < 0.001) activation of NF-κB through TLR5 than equimolar FlaB protein (Fig. 1C). Therefore, the Derp2-FlaB fusion protein was used in later experiments to test its protective effects in the Derp2 allergen-induced asthma model.
The Derp2-FlaB fusion protein effectively suppresses airway hyperreactivity
Noninvasive AHR was measured 24 hours after the final Derp2 challenge. For the baseline Penh readings, mice were challenged with control PBS aerosol and administered increasing concentrations of methacholine (6.25, 12.5, 25 and 50 mg/mL). The Derp2 group showed enhanced AHR to the bronchoconstrictor methacholine, but the Penh values significantly decreased when sensitized mice were treated with the Derp2-FlaB fusion protein, showing Penh values similar to those of the negative control group (Fig. 3). Mice in the Derp2 + FlaB group showed lower Penh values (not significant) than those of the Derp2 group, but significantly higher (P < 0.05) values than those of the Derp2-FlaB fusion protein group (Fig. 3). The FlaB adjuvant group, without Derp2 allergen, showed higher Penh values than the PBS and Derp2-FlaB fusion protein groups, but the values were much lower than those of the Derp2 group (Fig. 3). This result explains the superior effects of the Derp2-FlaB fusion protein in suppressing airway hyperactivity.
Fig. 3
Assessment of airway hyperresponsiveness following allergen sensitization and challenge. Airway responsiveness to methacholine was determined in the PBS, Derp2, Derp2 + FlaB and Derp2-FlaB groups (n = 5 for each). The data are expressed as the mean ± SEM.
PBS, phosphate-buffered saline; FlaB, flagellin B; SEM, standard error of mean; ns, not significant.
*P < 0.05, †P < 0.01.

The Derp2-FlaB fusion protein effectively suppresses airway inflammation
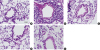
Lung histology was analyzed to investigate the efficacy of the FlaB-Derp2 vaccine in modulating airway inflammation. Typical pathological features of asthma were observed in the Derp2 group, including the infiltration of numerous inflammatory cells around the bronchioles, alveoli and blood vessels (Fig. 4B). However, mice treated with the Derp2-FlaB fusion protein showed a marked decrease in inflammatory cell infiltration, and their lung tissues showed almost normal histology that was comparable to that of the PBS control group (Fig. 4A and D). We found that Derp2 + FlaB treatment also reduced inflammatory cell infiltration in the lungs compared with the Derp2 group, but was less effective than the Derp2-FlaB fusion protein treatment (Fig. 4C and D). The FlaB adjuvant group showed less infiltration of inflammatory cells than the Derp2 group (Fig. 4E).
Fig. 4
Histological evaluation of lung tissues. Lung tissues were fixed with 10% formalin. Lung sections were stained with H&E and observed by optical microscopy (× 200). Representative images were obtained from the histology sections from the (A) PBS, (B) Derp2, (C) Derp2 + FlaB, (D) Derp2-FlaB and (E) FlaB groups.
H&E, hematoxylin and eosin; PBS, phosphate-buffered saline; FlaB, flagellin B.
The Derp2-FlaB fusion protein effectively suppresses Derp2-specific IgE in the serum
After the final allergen challenge, we collected serum from the mice and measured Derp2-specific IgE by ELISA. Derp2-specific serum IgE was significantly decreased (P < 0.001) in mice treated with the Derp2-FlaB fusion protein compared with the Derp2 group (Fig. 5). Derp2-specific serum IgE in the Derp2 + FlaB mixture-treated mice was also significantly lower (P < 0.01) than in the Derp2-treated mice, but slightly higher than in the Derp2-FlaB fusion-treated mice, although statistical significance was not observed (Fig. 5). The FlaB group showed a comparable level of Derp2-specific serum IgE to that of the Derp2 group (Fig. 5). These data indicate that the Derp2-FlaB fusion protein more effectively suppressed Derp2-specific serum IgE production than the Derp2 + FlaB mixture.
Fig. 5
Derp2-specific serum IgE levels in tested mice determined by ELISA. The data are expressed as the mean ± SEM.
IgE, immunoglobulin E; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of mean; FlaB, flagellin B; PBS, phosphate-buffered saline; ns, not significant.
*P < 0.01, †P < 0.001.

The Derp2-FlaB fusion protein effectively suppresses eosinophil infiltration and Th2 cytokine production in BAL fluid
We further assessed inflammatory cell types recovered in BAL fluid. Total cell numbers recovered in BAL fluid were high in mice treated only with Derp2, but significantly reduced in Derp2 + FlaB-treated mice and further reduced when mice were treated with the Derp2-FlaB fusion protein (Fig. 6A). By morphometric analysis, treatment with the Derp2-FlaB fusion protein significantly reduced (P < 0.05) the number of eosinophils in BAL fluid compared with the Derp2 group. Treatment with Derp2 + FlaB also significantly reduced (P < 0.05) the number of eosinophils, although eosinophil infiltration was significantly higher (P < 0.01) in this group than in the Derp2-FlaB fusion-treated animals (Fig. 6A). Mice treated with only the FlaB adjuvant showed low eosinophil infiltration (Fig. 6A). IL-4, IL-5 and IL-13 in BAL fluid were significantly increased in Derp2-treated mice, but significantly decreased when mice were treated with the Derp2 + FlaB mixture or the Derp2-FlaB fusion protein (Fig. 6B-D). Treatment with Derp2 + FlaB or Derp2-FlaB did not cause a significant difference in the level of IL-4, IL-5 or IL-13 in BAL fluid (Fig. 6B-D). The FlaB adjuvant group showed low IL-4, IL-5 and IL-13 in BAL fluid (Fig. 6B-D). These results suggest that the intranasal administration of Derp2 + FlaB or Derp2-FlaB fusion protein inhibits allergic airway inflammation by inhibiting Th2-immune responses.
Fig. 6
Inflammatory cell and cytokine analysis of the BAL fluid. (A) BAL cells. BAL was carried out 24 hours after methacholine challenge in the mice. Cytokines IL-4 (B), IL-5 (C) and IL-13 (D) in BAL fluid were determined by ELISA. The data are expressed as the mean ± SEM.
BAL, bronchoalveolar lavage; IL, interleukin; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of mean; FlaB, flagellin B; PBS, phosphate-buffered saline; ns, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.

DISCUSSION
We have previously demonstrated that a therapeutic dose of flagellin was an excellent tolerogenic adjuvant inhibiting allergic asthma in OVA- and HDM-induced asthma mouse models.9 In the present study, we compared the therapeutic role of the Derp2-FlaB fusion protein with that of a mixture of Derp2 and FlaB in a Derp2-induced asthma mice model. We found that the effect of the Derp2-FlaB fusion protein was superior to that of the mixture of Derp2 and FlaB in relieving allergic asthma, as evidenced by reductions in AHR, lung inflammation, eosinophil infiltration, Derp2-specific IgE and Th2-cytokine production. The fusion protein should be easier to translate into clinical studies, since it will be regarded as a single medicinal entity and would require simpler preclinical and clinical verifications than mixture formulations that must undergo separate safety and efficacy trials.
Antigens fused with flagellin are usually more effective in stimulating protective immunity through more robust induction of TLR5 signaling. A fusion protein of recombinant FlaB and pneumococcal surface protein A (PspA) was able to elicit more efficient protective mucosal immune responses against pneumococcal infection than PspA alone or a stoichiometric mixture of PspA and FlaB.15 Similar enhanced immune responses against partner antigen were observed with the FlaB-tetanus toxin fragment C (TTFC) fusion.16 A fusion protein of flagellin and OVA showed a TLR-mediated immunomodulating capacity that was superior to that of a mixture of both proteins, leading to the prevention of intestinal allergy in mice.30 In this study, we showed that the Derp2-FlaB fusion protein more effectively stimulated TLR5 signaling than FlaB alone at equimolar concentrations, while the FlaB-Derp2 fusion protein stimulated less TLR5 signaling than FlaB (Fig. 1C). From our past experience, the stability and conformational characteristics of fusion proteins seem to be responsible for the differences between the biological activities of those fusion proteins.1516 Flagellins are self-assembling and properly hide their TLR5-stimulating domains,3132 and a fusion-partner polypeptide may either interfere with or promote the polymer formation of flagellins, rendering the TLR5-interacting domains more exposed or more hidden. We speculate that the fusion of Derp2 to the N-terminus of FlaB might have interfered with FlaB polymer formation to expose more TLR5-binding domains. Thus, the Derp2-FlaB conformation more potently enhanced TLR5 signaling and displayed a more efficacious immunomodulatory capacity in Derp2-induced allergic asthma than the Derp2 + FlaB mixture.
In this study, we applied a widely used asthma model in which BALB/c mice were sensitized 3 times at 1-week intervals with Derp2 and HDM plus alum and 7 daily intranasal Derp2 challenges.25 We started treatment 4 days after the first sensitization with 3 weekly intranasal administrations, while previous studies performed treatment prior to sensitization. Since most patients are already exposed to target allergens, we suggest that our experimental model reproduces rather real clinical situations requiring allergen-specific immunotherapy (SIT). Sensitized mice treated with Derp2 protein alone developed severe allergic airway inflammation, the extensive infiltration of inflammatory cells, such as eosinophils, and high levels of allergen-specific serum IgE. The intranasal co-treatment of sensitized mice with Derp2 and FlaB, especially as the Derp2-FlaB fusion protein, presented a dramatic alleviation of airway inflammation. We found that sensitized mice treated with FlaB alone developed less severe allergic airway inflammation but higher levels of allergen-specific serum IgE than those of the Derp2 group. This interesting result drove us to hypothesize the difference in routes for sensitization and treatment would have contributed to this result. Sensitization with HDM and Derp2 was carried out through the intraperitoneal route, which should have induced high levels of serum IgE. On the other hand, FlaB treatment was done through the intranasal route. The intranasal FlaB-only treatment might not be sufficient to suppress IgE responses in the systemic compartment, while local airway allergic reactions were significantly suppressed by the mucosal FlaB treatment. In our previous study, we have shown that ex vivo FlaB-only treatment of patients-derived DCs was sufficiently efficacious in inducing Treg cells from co-cultured autologous peripheral lymphocytes.9 Combination of FlaB with Derp2 allergen, either in combination or as a physical fusion protein, should have been more efficacious and caused IgE suppression even in the serum/systemic compartment. As proposed in our previous FlaB-PspA fusion study,15 stimulation of the same antigen presenting cell with both antigen/allergen and adjuvant (FlaB) should be significantly more potent in inducing desired therapeutic effects than the mixture formulation of the 2 components. Repa et al.33 also evaluated the influence of the sensitization route on the development of IgE antibody responses and allergic airway inflammation using a mouse model of type I allergy to the major birch pollen allergen Bet v 1. They had shown that aerosol sensitization resulting in the lowest systemic IgE but strong airway inflammation, while IP sensitization resulting in the lowest airway inflammation but higher IgE than aerosol sensitization. These findings indicate that the intranasal treatment of Derp2 protein might have further stimulated allergic airway inflammation, while co-treatment with FlaB adjuvant mediated the suppression of the Derp2 allergen effect.
When we treated the sensitized mice with the same dose of Derp2 + FlaB or Derp2-FlaB fusion protein via the sublingual route, we did not observe an obvious inhibitory effect on airway inflammation (data not shown). Flagellin exhibits effective mucosal adjuvant activity: immunization through the intranasal and sublingual routes induces considerable systemic and local antibody responses.83435 However, as the intranasal route is superior to the sublingual route in inducing systemic immune responses, the administration route may affect the immunomodulatory threshold of flagellin in inducing the suppressive immunomodulation of airway inflammation. The reason why this occurred should be further studied, given that the sublingual route is the more preferred site for SIT for allergic asthma.
Asthma is characterized in large part by an abnormally polarized Th2-type immune response. Asthma immunotherapy should constitute immune intervention driving allergen-specific Th2 responses to a protective or less harmful Th1 response or generating Tregs that induce anergy or tolerance of allergen-specific lymphocytes. Flagellin is capable of differentially affecting Th1 or Th2 immune responses, depending on the experimental model, co-administered antigen(s) and flagellin dosage.363738 The immunomodulatory activity of flagellin has been reported to ensue primarily through DCs.394041 Our previous studies showed that treatment with 0.025 µg of FlaB increased AHR, airway inflammation and IL-13 in the BALF, while a high dose of FlaB (10 µg) ameliorated OVA-induced or HDM-induced asthma by inhibiting Th1/Th2/Th17 responses in a TLR5-dependent manner by generating rDCs and Treg cells.9 In this study, to ensure the use of equal molar concentrations, we treated sensitized mice with 12 µg of FlaB and 4 µg of Derp2 or 16 µg of Derp2-FlaB fusion protein, which was a slightly higher dose of FlaB than we previously used.9 Accordingly, we hypothesize that the mechanism underlying the Derp2-FlaB fusion protein-mediated suppression of Derp2-induced allergic asthma is the same as what was observed in our previous studies as evidenced by similar cytokine and IgE production profiles.
In conclusion, we report an allergen-specific immunomodulatory role for an allergen-flagellin fusion protein using a mouse asthma model. Our observations suggest that the Derp2-flagellin fusion further potentiates the allergen-specific immunomodulatory properties of flagellin compared with simply mixing it with allergens. The Derp2-FlaB fusion protein is a promising vaccine candidate for allergic asthma SIT.
 XML Download
XML Download