

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Lipopolysaccharides (LPS) known as lipoglycans and endotoxin, is an important component of the Gram-negative bacteria outer membrane (1). It is involved in a variety of immune responses by binding CD14/TLR4/MD2 receptor complexes in many cell types including monocytes, dendritic cells, macrophages, and B cells (23). It has been shown that LPS stimulates innate immunity and prolonged exposure to high doses of LPS induces oxidative stress caused by production of reactive oxygen species (ROS) (34). Since ROS molecules are highly reactive low molecular weight lipophilic species, the ROS molecules can easily penetrate cell walls and induce cell damage (35). Therefore, high level of ROS production not only induces cell damage but also stimulates downstream signaling pathways and expends the immune responses (46).
The ROS, such as superoxide anion (O2−), hydroxyl (OH−), perosyl (ROO−) or hydrogen peroxide (H2O2), are mainly produced in the mitochondria during cellular metabolism through the respiratory chain (78). These ROS are highly unstable and reactive, so they can easily react with damaged cell compartments such as lipids, proteins, and DNA (9). On the other hand, the cells have a defense system with antioxidant enzymes including superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) against harmful ROS (10). For example, SOD converts O2− anions to H2O2 and H2O2 is rapidly removed by detoxifying enzymes such as GPx. In addition, CAT decomposes and removes H2O2 into water and oxygen. Therefore, antioxidant enzymerich bioactive compounds could protect the body from chronic diseases caused by ROS.
Recent studies have suggested that induction of detoxifying enzyme activity is regulated by transcriptional levels in the cis-acting element located within the promoters of the detoxifying enzyme genes called the antioxidant-responsive element (ARE) (11). This element is also found in the gene promotors encoding cellular antioxidant enzymes, such as haem oxygenase-1 (12) and glutathione-S-transferase (13) and is activated by various oxidative stress. Therefore, binding to DNA site of ARE is one of the major mechanisms in cellular defense from ROS, resulting in target gene expression associated with detoxification and elimination of ROS (14).
Ginseng is an herb that is famous in eastern Asian countries and has recently been used widely in Western countries. One of the components of ginseng, ginsenoside, has been reported to have various biological effects including anti-tumor, anti-inflammation and anti-stress activities (151617). The ginsenosides were classified into two groups, protopanaxadiol (PPD) and protopanaxatriol (PPT) (18). The difference between PPD and PPT types is that PPD types have carboxyl group at the C-6 position, but PPT types do not. The PPD group includes ginsenosides of Rb1, Rb2, Rc, Rd, Rg3 and Rh2, and the PPT group includes ginsenosides of Re, Rf, Rg1, Rg2 and Rh1. Among the PPT type ginsenosides, Rh1 and Rg2 can be classified as minor ginsenosides, which are deglycosylated from the major ginsenosides. It has been reported that ginsenosides Rh1 and Rg2 protect the rheological functions of erythrocytes against oxidative stress (19). However, the effects of Rh1 and Rg2 on antioxidant activity and their regulatory effects on the antioxidant enzymes have not been studied. Therefore, we investigated the effect of ginsenosides Rh1 and Rg2 on antioxidant activity and antioxidant-related gene expression in LPS-induced RAW 264.7 cells.
MATERIALS AND METHODS
Reagents
3-(4, 5-dimethylthiazole-2-yl)-2, 5-diphenyltetrazolium bromide (MTT, #M6494) was purchased from Invitrogen (Waltham, MA, USA). Dulbecco's modified Eagle's medium (DMEM, #11963-092), and fetal bovine serum (FBS, #10082147) were purchased from Gibco (Waltham, MA, USA). 1, 1-Diphenyl-2picrylhydrazyl (DPPH, #D9132), lipopolysaccharide (LPS, E. coli, O111:B4, #L2630) and 2′-7′-Dichlorofluorescin diacetate (DCF-DA, #D6883) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Phosphate buffered saline (PBS, #EBA-1105) and reveres transcription 5X master mix (#EBT-1511) were purchased from ELPIS-BIOTECH (Daejeon, Korea). Tri-RNA reagent (#FATRR-001) was purchased from Favorgen (PingTung, China). Ginsenosides Rh1 and Rg2 (purity 99%) were purchased from Ace EMzyme (Anseong-si, Korea). Rh1 and Rg2 is mixed in a 1:1 ratio.
Cell culture and treatment
The murine macrophage RAW 264.7 cell was purchased from Korea Cell Line Bank (KCLB, #40071, Seoul, Korea) and maintained in DMEM supplemented with 10% FBS, 1% penicillin/streptomycin at 37℃ in a humidified atmosphere with 5% CO2 (HERAcell 150i, Thermo Electron Corp., Waltham, MA, USA). For experiments, RAW 264.7 cells were seeded in 96-well plates at the density of 1 × 104 cells/well. After serum starvation for 24 h, cells were treated with FBS-free medium containing various concentrations (0, 1, 5, 10, 25 or 50 µg/ml) of ginsenosides Rh1 and Rg2 followed by treatment with LPS (500 ng/ml) for indicated time in each Figure legend.
Free radical-scavenging activity of DPPH
The DPPH radical scavenging activity of the Rh1 and Rg2 was determined according to the previous report (20). Briefly, samples at various concentrations were added to 5 µl of DPPH (2.5 mg/ml) in 100% ethanol. The mixture was mixed and allowed to stand for 30 mins at 25℃ dark conditions. The absorbance was measured at 517 nm using microplate reader (TECAN, Männedorf, Switzerland). Vitamin C (Vit. C) was used as a positive control. The free radical-scavenging capacity was expressed by relative percentage of control.
ABTS analysis
The ABTS [2,2′-azino-bis(3-ethlybenzthizoline-6-sulphonic acid)] assay was performed to scavenge the ABTS radical cation in comparison to the compounds. The radical cation was prepared by mixing 7 mM ABTS with 2.4 mM potassium persulfate (1:1 v/v) and leaving the mixture for 24 h until the reaction was completed and the absorbance was stable. The ABTS radical solution was diluted with PBS and absorbance of 0.7 (±0.02) at 732 nm. The assay was conducted with diluted ABTS radical solution mixed with samples, and the measurements were taken at 732 nm after 30 mins. The anti-oxidative activity of the samples was calculated by determining the decrease in absorbance.
MTT cell viability assay
The mitochondrial-dependent reduction of MTT to formazan was used to measure cell respiration as an indicator of cell viability (21). Briefly, RAW 264.7 cells were seeded in 96-well plates at the density of 1 × 104 cells/well. After 24 h of incubation, fresh FBS free medium were replaced for 24 h. The adhered cells were treated with FBS-free medium containing various concentrations (0, 1, 5, 10, 25 or 50 µg/ml) of the compound. After 24 h, MTT (12 mM) was prepared in FBS-free DMEM media to give a total volume of 100 µl in each well and the cells were incubated for 2 h at 37℃ and 5% CO2. The medium was the removed and the formazan precipitate was solubilized in DMSO. The absorbance was measured at 540 nm on a microplate reader (TECAN).
Intracellular ROS scavenging activity
Intracellular ROS levels were measured by detecting the fluorescent intensity of cells with 2′,7′-dichlorodihydrofluoresceindiacetate (DCFH-DA) (22). RAW 264.7 cells were seeded in 96-well plate at the 1 × 104 cells/well. Cells were then starved for 24 h in the serum free DMEM. Then the cells were treated with various concentrations of compounds. After 1 h, the cells were stimulated with or without medium containing LPS (500 ng/ml) at 37℃ for 24 h. Cells were washed with PBS and incubated at 37℃ for 30 min (in dark) with the probe at a final concentration of 10 µM DCFH-DA. Cells were washed with PBS and the fluorescence intensity of DCF was measured by using the fluorescence microplate reader with an excitation wavelength of 485 nm and an emission wavelength of 530 nm (TECAN).
Plasmids and luciferase assay
The DNA construct of antioxidant response element (ARE) encoding pARE-luc plasmid was obtained by Promega (Madison, WI, USA). The DNA construct contains four copies of ARE (5′-TGACnnnGCA-3′) that drives transcription of the luciferase reporter gene luc2P (Photinus pyralis). RAW 264.7 cells were plated on 12-well plates at 5 × 104 cells/well. For a luciferase assay, cells were transfected in Opti-MEM (#31985-088, Invitrogen) with lipofectamine mixture containing the pARE-luc reporter plasmid along with pRL-CMV for 18 h. Cells were pretreated with 10 µg/ml of Rh1 and Rg2 compounds for 1 h followed by treatment with 500 ng/ml of LPS for 6 h. The cells were collected after treatment, and the luciferase activity was assayed with the dual luciferase kit (E1960, Promega) using a TD-20/20 luminometer (Turner Designs, San Jose, CA, USA). Since pRL-tk contains the Renilla luciferase gene, the expression and transfection efficiencies were normalized with the Renilla luciferase activity. Transfections were performed in triplicate, and each experiment was repeated at least three times.
Real-time reverse transcription polymerase chain analysis
Quantitative RT-PCR (qRT-PCR) assay was used to analyze the mRNA expression of anti-oxidative enzymes including Cu/ZnSOD, MnSOD, catalase, or GPx according to the previous report (23). Total RNA was isolated from cells using Tri-RNA reagent according to the manufacturer's instructions. The 1 µg of total RNA in each group was reverse transcribed into single-stranded cDNA using the reverse transcription 5X master mix. The mRNA expression of the genes was quantified with iQ™ SYBR green supermix (#170-8882, Bio-Rad, Hercules, CA, USA) using a CFX Connect™ (Bio-Rad). The relative gene expression was calculated using the 2−ΔΔct method, and GAPDH was used for normalization. All primer sequences used in qRT-PCR experiments are listed in Table 1.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 5 (version 5.02, GraphPad Software Inc., San Diego, CA, USA). One-way analysis of variance (ANOVA) followed by a Bonferroni multiple comparison was performed. A p value <0.05 was considered significant. All experiments were expressed as the mean ± SEM and were performed independently at least 3 times.
RESULTS
The effect of minor ginsenosides Rh1 and Rg2 on cell cytotoxicity of RAW 264.7 cells
Cell viability tests were performed using MTT assay to determine the experimental concentration of the compounds. The serum starved RAW 264.7 cells were treated with various concentrations of Rh1 and Rg2 compounds (1, 5, 10, 20, 40, 60, 80 and 100 µg/ml) for 24 h. The results showed that there was no cytotoxic effect on RAW 264.7 cells at the tested concentrations (Fig. 1). Therefore, ginsenosides Rh1 and Rg2 concentrations of 1 to 50 µg/ml were selected in subsequent experiments.
Antioxidant capacities of minor ginsenosides Rh1 and Rg2
The radical scavenging capacity of the compounds were determined by DPPH and ABTS assay. Various concentrations of Rh1 and Rg2 compounds were mixed with the free radical of DPPH solution and then residual free radical DPPH was measured at 518 nm using a plate reader. Surprisingly, the scavenging effect of DPPH radical was not shown by treatment with any concentrations of Rh1 and Rg2 compounds (Fig. 2A). The in vitro radical scavenging effect of the Rh1 and Rg2 compounds was confirmed with ABTS analysis. Consistent with the data of the DPPH assay, the reduction of ABTS radical was also not observed by treatment of the Rh1 and Rg2 compounds (Fig. 2B).
Minor ginsenosides Rh1 and Rg2 inhibit LPS-induced intracellular ROS production
To investigate the effect of Rh1 and Rg2 on intracellular ROS production, RAW 264.7 cells were treated with various concentrations of the compounds for 24 h followed by treatment with serum-free medium containing H2DCF-DA. Interestingly, Rh1 and Rg2 compound treatment inhibits intracellular ROS production in a dose-dependent manner in unstimulated RAW 264.7 cells (Fig. 3A). When the cells were treated with 500 ng/ml LPS for 2 h, the ROS levels were strongly increased by more than 8-fold compared to the unstimulated condition (Fig. 3B). When cells were co-treated with Rh1 and Rg2 compounds following LPS treatment, LPS-induced ROS levels were significantly suppressed at the concentrations of 10, 25, and 50 µg/ml (Fig. 3B). Vitamin C (Vit C) was used as a positive control.
Minor ginsenosides Rh1 and Rg2 increase anti-oxidant activity
To investigate how ginsenosides Rh1 and Rg2 exert antioxidative effect by eliminating intracellular ROS production, pARE-luc was used to confirm that ginsenosides Rh1 and Rg2 regulates cis-acting element placed in the promoters of the detoxifying enzyme genes. For experiment, cells were transfected with pARE-luc to analyze the activity of the firefly luciferase reporter containing sequences of the anti-oxidant response elements and cotransfected with pRL-CMV to normalized reporter activity with the Renilla luciferase expression. Treatment with 500 ng/ml LPS significantly decreased ARE transcriptional activity whereas treatment with Rh1 and Rg2 reversed LPS-induced inhibition (Fig. 4).
Minor ginsenosides Rh1 and Rg2 induce anti-oxidant enzymes in RAW 264.7 cells
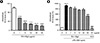
To confirm how Rh1 and Rg2 compounds inhibit LPS-induced intracellular ROS production, mRNA levels of anti-oxidant enzymes, such as Cu/Zn SOD, Mn-SOD, CAT, and GPx were examined using qRT-PCR analysis. As shown in Fig. 5, treatment with Rh1 and Rg2 compounds significantly increases the mRNA expression of Cu/Zn SOD, Mn-SOD, CAT, and GPx compared to that in the LPS-induced control group at 10, 25, and 50 µg/ml.
DISCUSSION
Ginsenosides Rh1 and Rg2 have been reported to possess various biological activates, including anti-inflammatory, anti-allergic and antioxidant properties (151617). In addition, recent report shows that ginsenosides Rh1 and Rg2 have an anti-rheological function of erythrocytes against oxidative stress (19). However, there was lack of information on the regulatory effect of ginsenosides Rh1 and Rg2 on antioxidant activity and antioxidant enzymes. In this study, we aimed to investigate the effect of Rh1 and Rg2 on antioxidant activity and antioxidant-related gene expression in LPS-stimulated RAW 264.7 cells.
Many studies have shown the antioxidant effects of bio-active compounds using DPPH and ABTS assay (2425). Both DPPH and ABTS are free radicals and if the compound itself is effective as an antioxidant, the compound can remove free radicals from the tube reaction. Ginsenosides Rh1 and Rg2 did not show in vitro radical scavenging effects on DPPH and ABTS radicals (Fig. 2). Instead, these bio-active compounds showed an inhibitory effect on intracellular ROS production in RAW 264.7 cells (Fig. 3). Cells treated with high doses of LPS (500 ng/ml) for 2 h strongly enhanced intracellular ROS levels, whereas treatment with Rh1 and Rg2 significantly inhibited LPS effect. We therefore investigated how ginsenosides Rh1 and Rg2 inhibited LPS-stimulated intracellular ROS production.
Previous studies have suggested that antioxidant defensive genes contribute to the response to cellular oxidative stress along with induction of genes encoding catalytic and modifier subunits, which are important for glutathione synthesis (26). These antioxidant defensive genes are up-regulated through the ARE located within the promoters of their respective genes, such as HO-1 (12) or glutathione-S-transferase (13). Interestingly, treatment with LPS reduced ARE reporter activity, whereas treatment with ginsenosides Rh1 and Rg2 reversed LPS-induced inhibition (Fig. 4). Therefore, it is possible that ginsenosides Rh1 and Rg2 have antioxidant effects in RAW-264.7 cells by the upregulation of ARE reporter activity. Recent report has demonstrated that the enhancement of HO-1 expression inhibits LPS-induced macrophage activation through upregulation of the ARE binding site of the HO-1 promoter and interaction with nuclear factor erythroid-2-related factor (Nrf-2) (27). To understand the mechanism of ginsenosides Rh1 and Rg2 having an antioxidative effect through the regulation of ARE-luc activity, investigating the effects of ginsenosides Rh1 and Rg2 on HO-1 expression, Nrf-2 transcriptional activity, and interaction between HO-1 and Nrf-2 will be good targets for future study.
It has been well established that SODs, such as Cu/Zn-SOD and Mn-SOD, are the first line of defense for superoxide free radicals (28). SODs have distinct subcellular localization depending on the metal ion cofactors (29). For example, Cu/Zn-SOD is mainly localized in the cytoplasm and Mn-SOD is localized in the mitochondria (29). Figs. 5A and B show that treatment with LPS inhibits all mRNA levels of SOD, but treatment with ginsenosides Rh1 and Rg2 results in dose-dependent increase in the mRNA levels of SOD. It suggests that ginsenosides Rh1 and Rg2 strongly upregulate expression of SOD in cytoplasm and mitochondria. In addition to SOD, catalase and GPx were found in many subcellular components including peroxisomes and cytoplasm, and mitochondria and nucleus, respectively. Interestingly, Rh1 and Rg2 also reversed these antioxidant enzymes from LPS-induced inhibition (Figs. 5C and C). Together, our current study found inhibitory effect of ginsenosides Rh1 and Rg2 on intracellular ROS production through regulation of pARE-luc promoter activity and upregulation of antioxidant enzyme expressions. It suggests that ginsenosides Rh1 and Rg2 would be good natural agents for treating various human diseases caused by oxidative stress with antioxidant effect.





 XML Download
XML Download