

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Native turnip yellow mosaic virus (TYMV) is relatively unreactive to maleimide agents, indicating few reactive thiol groups on TYMV. In the present study, we aimed to construct TYMV mutants that have reactive cysteine residues on the surface. To this end, we prepared a library of TYMV mutants where the Thr residue at the C-terminus of coat protein (CP) was replaced by a random sequence of six amino acids that included one cysteine. This library was introduced into Nicotiana benthamiana by agroinfiltration. The CP sequence of the TYMV RNA isolated from inoculated leaves was amplified by reverse transcription-PCR and then used to construct a second library. This process was repeated one more time, and the CP sequences of the TYMV RNA in the inoculated leaves were analyzed. Based on the analysis of over 11,000 CP sequences, the Cys mutants representing most abundant TYMV RNAs were constructed. Analysis of the mutants showed that four Cys mutants were nearly comparable to wildtype with respect to CP and viral RNA levels in N. benthamiana. All these mutants were highly reactive to fluoresceine-5-maleimide. This demonstrates that TYMV can be modified to have additional functional groups on the surface that would be useful for drug delivery.
Go to : 
REFERENCES
1). Dreher TW. Turnip yellow mosaic virus: transfer RNA mimicry, chloroplasts and a C-rich genome. Mol Plant Pathol. 2004; 5:367–75.

2). Steinmetz NF, Evans DJ. Utilization of plant viruses in bionanotechnology. Org Biomol Chem. 2007; 5:2891–902.
3). Yildiz I, Shukla S, Steinmetz NF. Applications of viral nanoparticles in medicine. Curr Opin Biotechnol. 2011; 22:901–8.
4). Wang Q, Lin T, Tang L, Johnson JE, Finn MG. Icosahedral virus particles as addressable nanoscale building blocks. Angew Chem Int Endl. 2002; 41:459–62.
5). Steinmetz NF. Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomedicine. 2010; 6:634–41.
6). Barnhill HN, Reuther R, Ferguson PL, Dreher T, Wang Q. Turnip yellow mosaic virus as a chemoaddressable bionanoparticles. Bioconjug Chem. 2007; 18:852–9.
7). Michels B, Leimkühler M, Lechner MD, Adrian M, Lorber B, Witz J. Polymorphism of turnip yellow mosaic virus empty shells and evidence for conformational changes occurring after release of the viral RNA. A differential scanning calorimetric study. Eur J Biochem. 1999; 264:965–72.
8). Katouzian-Safadi M, Berthet-Colominas C. Evidence for the presence of a hole in the capsid of turnip yellow mosaic virus after RNA release by freezing and thawing. Decapsidation of turnip yellow mosaic virus in vitro. Eur J Biochem. 1983; 137:47–53.
9). Kim D, Lee Y, Dreher TW, Cho TJ. Empty Turnip yellow mosaic virus capsids as delivery vehicles to mammalian cells. Virus Res. 2018; 252:13–21.
10). Rohovie MJ, Nagasawa M, Swartz JR. Virus-like particles: Next-generation particles for targeted therapeutic delivery. Bioeng Transl Med. 2017; 2:43–57.
11). Schoonen L, van Hest JC. Functionalization of protein-based nanocages for drug delivery applications. Nanoscale. 2014; 6:7124–41.
12). Shin HI, Cho TJ. A sequence in coat protein open reading frame is required for Turnip yellow mosaic virus replication. J Bacteriol Virol. 2011; 41:109–16.
13). Cho TJ, Dreher TW. Encapsidation of genomic but not subgenomic Turnip yellow mosaic virus RNA by coat protein provided in trans. Virology. 2006; 356:126–35.
14). Canady MA, Larson SB, Day J, McPherson A. Crystal structure of turnip yellow mosaic virus. Nat Struct Biol. 1996; 3:771–81.
15). Shin HI, Chae KH, Cho TJ. Modification of Turnip yellow mosaic virus coat protein and its effect on virion assembly. BMB Rep. 2014; 46:495–500.
16). Bransom KL, Weiland JJ, Tsai CH, Dreher TW. Coding density of the turnip yellow mosaic virus genome: roles of the overlapping coat protein and p206-readthrough coding regions. Virology. 1995; 206:403–12.
17). Hipper C, Brault V, Ziegler-Graff V, Revers F. Viral and cellular factors involved in phloem transport of plant viruses. Front Plant Sci. 2013; 4:154.
18). Shin HI, Kim IC, Cho TJ. Replication and encapsidation of recombinant Turnip yellow mosaic virus RNA. BMB Rep. 2008; 41:739–44.
19). Ren Y, Wong SM, Lim LY. Folic acid-conjugated protein cages of a plant virus: a novel delivery platform for doxorubicin. Bioconjug Chem. 2007; 18:836–43.
20). Aljabali AA, Shukla S, Lomonossoff GP, Steinmetz NF, Evans DJ. CPMV-DOX delivers. Mol Pharm. 2013; 10:3–10.
Go to :
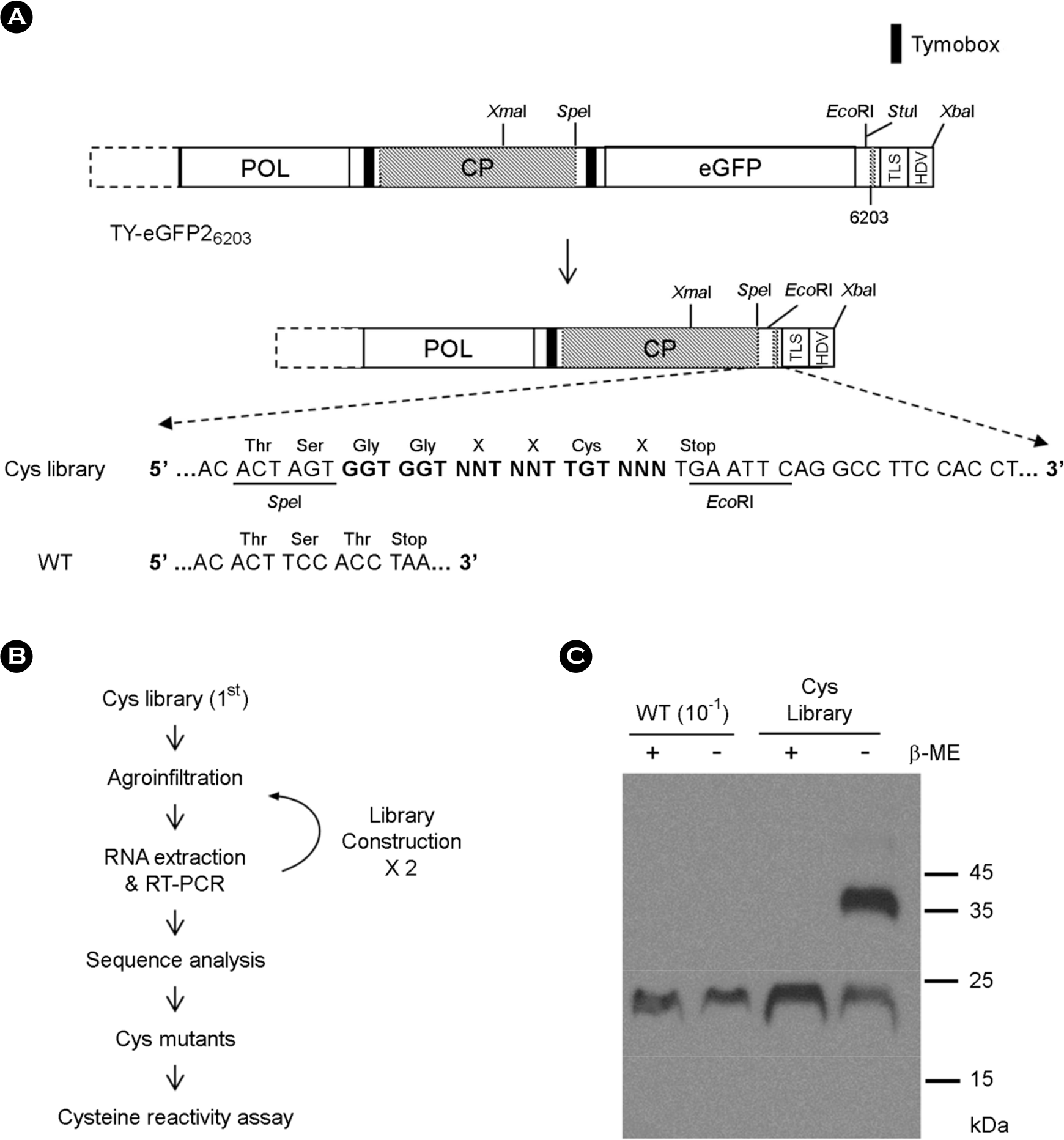 | Figure 1.TYMV Cys mutant library. (A) Library construction. A random sequence of three amino acids and one cysteine was introduced to the C-terminal part of TYMV CP by PCR-amplification and insertion of the PCR product into Xma I/ Eco RI sites of TY-eGFP26203 (for further details, see MATERIALS AND METHODS). The C-terminal extension is highlighted in bold face. (B) Screening of a Cys mutant library. A. tumefaciens containing the library was introduced to N. benthamiana. In a week, RNA was extracted from the inoculated leaf and the TYMV CP sequence was amplified by RT-PCR. The amplified DNA was used to generate the 2 nd library, which was introduced again to N. benthamiana. After this process was repeated, the TYMV CP sequence was examined by high-throughput sequencing analysis using encapsidated TYMV RNA. (C) Examination of CP. Leaf extract was prepared in the presence (+) or absence (-) of 20 mM β-mercaptoethanol (β-ME) from the leaf inoculated with the 3rd library or wild-type TYMV. 2 μl of each leaf extract sample (1:10 diluted sample for wildtype) were loaded onto 12.5% SDS-polyacrylamide gel. After electrophoresis, the proteins were examined by Western blot analysis using TYMV CP antibody. |
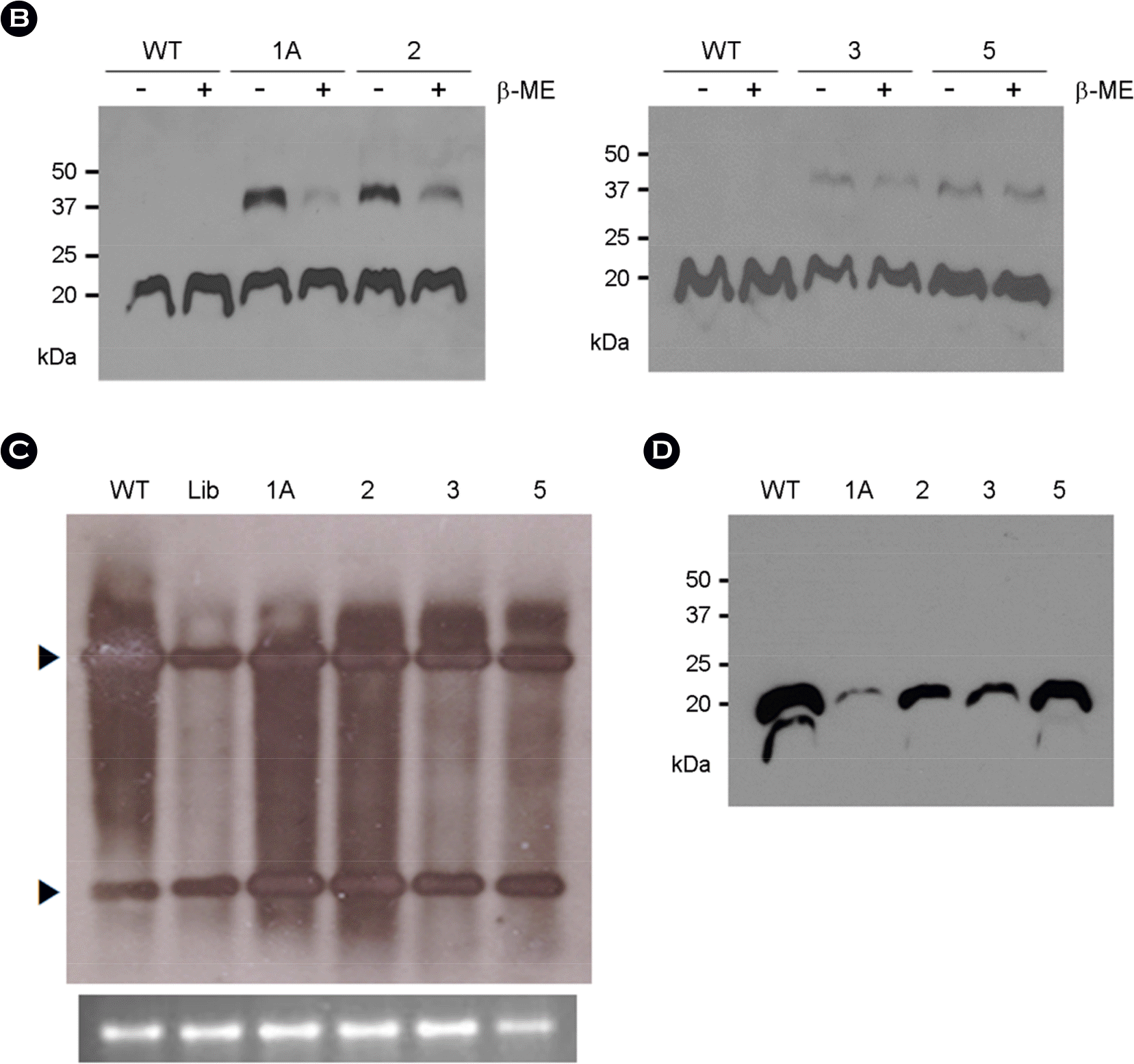 | Figure 2.Cys mutants. (A) Sequence of the Cys mutants. Nucleotide and amino acid sequences of Cys mutants are shown. In Cys1A, the nucleotide sequence upstream of the ACTAGT is GCC, whereas, in other mutants, the sequence is GAC. (B) Western blot analysis of CP in leaf extracts. 1 μg of total protein for each sample was loaded onto 12.5% SDS-polyacrylamide gel. After electrophoresis, the proteins were examined by Western blot analysis using TYMV CP antibody. (C) Northern blot analysis of TYMV RNA. 500 ng of total RNA were size-fractionated in a 1% agarose gel and examined by Northern blot analysis using the DIG-labeled TYMV CP DNA as a probe. The blots were developed by chemiluminescent immunodetection of DIG. Upper and lower arrowheads indicate genomic and subgenomic RNAs of TYMV, respectively. The panel below the Northern blot shows 25S rRNA stained with EtBr. (D) Systemic infectivity of the Cys mutants. Chinese cabbage was inoculated with the TYMV constructs containing wildtype or the Cys mutants, and young systemically infected leaf (5th leaf from the inoculated leaf) was collected two weeks post-inoculation. Total protein (1 μg) in the leaf extract prepared in the presence of β-ME was examined for its CP levels by Western analysis as described in Fig. 2B. |
 | Figure 3.Cysteine reactivity. Wildtype (WT) and Cys mutants were reacted with fluorescein-5-maleimide, and analyzed by SDS- PAGE. After viewing under a UV lamp (left panels), the gel was stained with Coomassie (right panels). Two different size markers were used, and some components (75 kDa and 25 kDa) of the marker used with Cys 1A and 5 were fluorescent. |
 XML Download
XML Download