

 PDF
PDF ePub
ePub Citation
Citation Print
Print


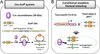
Cre-loxP System
Cre (Cre recombinase) is one of the tyrosine site-specific recombinases (T-SSRs) including flipase (Flp) and D6 specific recombinase (Dre) [12]. it was discovered as a 38-kDa DNA recombinase produced from cre (cyclization recombinase) gene of bacteriophage P1 [3456]. It recognizes the specific DNA fragment sequences called loxP (locus of x-over, P1) site and mediates site-specific deletion of DNA sequences between two loxP sites [78]. The loxP site is a 34 bp sequences consisting of a two 13 bp inverted and palindromic repeats and 8 bp core sequences (Figure 1A).
General principle of Cre-loxP system
Cre-loxP system is a widely used powerful technology for mammalian gene editing. This system has advantages which is very simple manipulation and do not require additional factors for efficient recombination [1]. Concerning the mechanism of Cre-loxP system, a single Cre recombinase recognizes two directly repeated loxP site, then the Cre excises the loxP flanked (floxed) DNA, thus creating two types of DNA with circular, excised and inactivated gene Y (Figure 1A). While the Cre-loxP system is predominantly used in genetic excision, it also induces the inversion and translocation of DNA between two loxP sites depending on the orientation and location of loxP sites [129].
To generate the spatiotemporally controlled mutant mice, two elements are needed in the Cre-loxP system [10111213]. First, Cre-driver strain is generated in which Cre recombinase is expressed by a promoter that specifically targets the cell or tissue of interest. Second, loxP flanked (floxed) DNA containing mouse strain is needed to be generated. Conditional knockout mice are then generated by breeding the Cre-driver strain with a floxed mouse strain (Figure 1B). The specificity and timing of recombination are controlled by used promoter and/or enhancer.
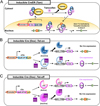
Inducible Cre-loxP system; CreERT
To achieve more accurate genetic functional studies and clinical applications using Cre-loxP system, it was required a more sophisticated technique that controls the Cre activation at the precise time and in a specific cell. An inducible Cre system is controlled by cell-specific regulatory elements (promoters and enhancers) and temporally inducible way by exogenous inducer such as tamoxifen (tam) or tetracycline (tet) [11014151617].
Tamoxifen-inducible Cre system is achieved by modified Cre protein fused with the estrogen receptor containing a mutated ligand binding domain (ER-LBD) [1415]. The fused Cre protein is called CreER recombinase, tamoxifen (also known as CreERT, Cre-ERT) and normally presented in the cytoplasm in a form that binds to heat shock protein 90 (HSP90). Upon binding the synthetic steroids (such tamoxifen or 4-hydoxytamoxifen; 4-OHT), the interaction is disrupted between HSP90 and CreERT. This causes nuclear translocation of CreERT and the interaction of Cre with loxP sites (Figure 2A). In this system, tamoxifen is systemically administered via intraperitoneal injection. To improve the function of targeted mutagenesis using CreERT, a new version of CreER, CreERT2 (Cre-ERT2), that is about ten-folds more sensitive to 4-OHT in vivo than CreERT has been generated [1518]. Thus, the use of CreERT2 is more preferred in several biological fields.
Another temporal and cell specific inducible system is tetracycline (Tet) system, also called doxycycline (Dox; a tetracycline derivative)-inducible Cre system. Dox is much more cost-effective, usable and efficient in controlling the Tet receptor (TetR) than tetracycline. This system is available in two modes, Tet-on and Tet-off, which permit Dox-dependent gene activation [1920] or inactivation [17]. The Tet systems consist of three elements, reverse tetracycline-controlled transactivator (rtTA), tetracycline-controlled transactivator (tTA) and tetracycline responsive element (TRE), also referred to as a tetracycline operon (TetO), which regulates a cre gene expression. When the rtTA binds to Dox, it can bind to the tetO7 (7 repeats of tetO) sequences and activates Cre gene expression (“Tet-on”) (Figure 2B), on the other hand, tTA binds to tetO7 sequences in normally, and when it is coupled with Dox, it can no longer bind to the tetO7 sequences and inactivates Cre expression (“Tet-off”) (Figure 2C). In Tet system, doxycycline is usually administered in feed or drinking water.
Tissue-specific promoters of Cre lines
Due to the usefulness of the Cre-loxP system, numerous organ-specific Cre and/or inducible Cre transgenic mouse lines have been created and are still being developed. Here, we provide some most commonly used tissue-specific promoter/enhancer of Cre driver lines in Table 1. The various organs were divided into 6 physiological systems: nervous, integumentary, immune, musculoskeletal, digestive and urogenital system. The useful promoters of Cre are selected and summarized based on International Mouse Phenotyping Consortium (IMPC), Mouse Genome Informatics (MGI), The Jackson Laboratory (JAX Cre repository) and National Center for Biotechnology Information (NCBI) databases.
Nervous system
Nervous system consists of two main parts, the central nervous system (CNS), which includes the brain and spinal cord, and the peripheral nervous system (PNS), which includes the all nerves of the entire body. Since the brain is very anatomically and functionally divided, we will divide it into three arbitrary regions; cerebrum, cerebellum and brain stem.
1) Cerebrum-specific promoters
The aldehyde dehydrogenase 1 family, member L1 (Aldh1L1) [21] and glial fibrillary acidic protein (Gfap) [2223] are expressed in astrocytes, but the GFAP is also known to be expressed in adult neural stem cells. Expression of calcium/calmodulin-dependent protein kinase II alpha (CaMK2α) [24] and glutamate receptor, ionotropic, kainate 4 (Grik4) [25] is detected in each CA1 and CA3 pyramidal neurons of hippocampus. Distal-less homeobox 1 (Dlx1) [26], distal-less homeobox 5/6 (Dlx5/6) [272829], glutamic acid decarboxylase 2 (Gad2), synthase 1, neuronal (nNos), parvalbumin (Pv, also called Pva, Parv) [30], somatostatin (Sst) and vasoactive intestinal polypeptide (Vip) promoters are predominantly expressed in inhibitory GABAergic interneurons [29], whereas the solute carrier family 17; sodium-dependent inorganic phosphate cotransporter, member 6 (Slc17a6) is expressed in excitatory glutamatergic, vesicular glutamate transport 2 (VGLUT2)-positive neurons [31]. The nestin (Nes) is also expressed in precursor cells of neuron and glia [32]. Platelet derived growth factor receptor, alpha polypeptide (Pdgfrα) is expressed in inner nuclear layer of the retina [33]. This promoter is useful in studies of retinal degenerative diseases such as age-related macular degeneration (AMD) and retinitis pigmentosa (RP). Expression of the leptin receptor (Lepr) is detected in hypothalamus and amygdala, and the promoter is used in research related to food intake and energy expenditure [34]. In the study of oligodendrocytes, proteolipid protein (myelin) 1 (PLP1) promoter is very useful in the studies of the gene function in myelination of mature oligodendrocytes [35]. Here, we provide the detailed expression regions of cerebral specific promoters in Table 2.
2) Cerebellum-specific promoters
In the cerebellum, Purkinje cells are the most salient cell type and play a major role in the cerebellar circuit. Purkinje cell protein 2 (Pcp2) is expressed in most Purkinje cells and is very useful for cerebellar studies [36].
3) Brainstem and Spinal cord-specific promoters
The brainstem is a region located between cerebrum and spinal cord and connects the two. Brainstem consists of three regions: midbrain, pons and medulla oblongata of the hindbrain. Solute carrier family 6; neurotransmitter transporter, dopamine, member 3 (Slc6a3 also known as DAT; dopamine transporter 1) is expressed in dopaminergic neurons of the midbrain and used for studying drug addiction and Parkinson's disease [37]. FEV; ETS oncogene family (Fev, also known as ePet) is expressed in the serotonin (5HT)-specific neurons of the midbrain, especially in the dorsal ad median raphe regions [38]. Neuropeptide Y receptor Y2 (Npy2r) expression is restricted in subsets of vagal sensory neurons in the lung and brainstem that regulate respiration [39]. In spinal cord, cadherin 3 (Cdh3) and 5-hydroxytryptamine; serotonin, receptor 6 (Htr6) are expressed in a subclass of inhibitory and excitatory interneurons in the dorsal horn of the spinal cord, respectively and these promoters have been used in neuronal functional studies [40].
4) PNS-specific promoters
Unlike the identification of enormous amounts of CNS-specific promoters of Cre, there are not many useful PNS-specific promoters. Actin-binding protein, Advillin (Avil)-driven Cre is expressed in all dorsal root ganglion (DRG) and trigeminal neurons but not in superior cervical ganglia (SCG) and is widely used for the studies of nociception in sensory neurons [4142434445].
Integumentary system
The integumentary system consists of the skin and its appendages such as hair, nails, sebaceous glands and sweat glands. Skin has at least three layers: epidermis (outer layer), dermis (middle layer) and hypodermis (deeper fatty layer, known as subcutaneous layer).
1) Skin and hair follicle-specific promoters
Epidermis, the top layer of skin, has five layers: stratum corneum, stratum lucidum, stratum granulosum, stratum spinosum and stratum basale. Each layer is composed of keratinocytes in different states [46]. Keratinocytes is the main cell type (~90%) of the epidermis and produces keratin, an intermediate filaments protein. Keratin 5 (Krt5) and Keratin 14 (Krt14) is expressed in basal keratinocytes of stratum basale [1847]. Keratin 10 (Krt10) is expressed in post-mitotic suprabasal keratinocytes [48]. Keratin 18 (Krt 18) expression is observed in Merkel cells of the basal epidermal layer [49]. Leucine-rich repeat-containing G protein-coupled receptor 6 (Lgr6, also known as GPCR) is expressed in hair placodes at embryonic stage and in follicle bulge of dermis at adult stage [50].
Immune system
Immune system is a host defense response against foreign invaders, and it protects the body from disease. This system consists of two working modes: innate immunity (also called non-specific, natural or native immunity), which is a first defense mechanism against invaders and initiates the adaptive immunity, and the adaptive immunity (also called specific or acquired immunity), which is systemic response mediating by lymphocytes such as T and B cells [515253]. The leukocytes, three major phagocytes (dendritic cells, macrophages and neutrophils), basophils, eosinophils, mast cells and natural killer cells (NK cells), are included in innate immunity and the helper T, cytotoxic T and B lymphocytes are included in adaptive immunity
1) Macrophage/Monocyte-specific promoters
Lysozyme 2 (Lyz2) is expressed in various myeloid cells including monocytes, macrophages and granulocytes. It is used for the studies of myeloid cell lineage tracing and the innate immunity [54].
2) Dendritic cells and Mast cells-specific promoter
Dendritic cells and mast cells are involved in innate immunity. To study the homeostasis and function of dendritic cells, Integrin alpha X (Itgax, also known as CD11c) promoter is used [55]. Chymase 1 (CMA1) is expressed in mast cells of tissues and can be used for the studying of their function or fate mapping [56].
3) T and B cells-specific promoters
T and B lymphocytes are representative immune cells involved in adaptive immunity. CD2 molecule (CD2) is expressed in peripheral T cells such as CD4 and CD8 cells [57]. The CD4 antigen (CD4), a membrane glycoprotein, is expressed in CD4-expressing T cells in lymphoid tissues and used for the studies of CD4-expressing T cell maturation and proliferation [5859]. Tumor necrosis factor receptor superfamily, member 4 (OX40, also known as ACT35, Txgp1, TXGP1L and CD134) is also expressed in activated CD4-positive T cells [60]. Expression of CD8 antigen, alpha chain (CD8a) is only observed in peripheral CD8-positive T cells but not CD4-T cells [61]. Forkhead box P3 (Foxp3) is used for studying the lineage stability of regulatory T cells (Treg) [62]. Lymphocyte protein tyrosine kinase (Lck) is mainly expressed in T cells and used for the selection and maturation studies of T cell development in thymus [6364]. CD19 antigen (CD19) is known to B-lymphocyte surface antigen and is expressed throughout B-lymphocyte development [6566].
Musculoskeletal system
The musculoskeletal system (also known as the locomoter system) is composed of two systems, muscular and skeletal system. This organ system is made up of the bones (the skeleton), muscles, cartilage, tendon, ligaments, joints and other connective tissues that bind tissues and organs together.
1) Mesenchymal stem cells/Osteochondro progenitors, Osteoblasts, Osteocytes and Osteoclasts-specific promoters
The bone formation (also known as ossification) process is caused by two types of ossification: intramembranous ossification (including skull and clavicle), bone is developed directly from mesenchyme or fibrous connective tissue and endochondral ossification, bone is created from a pre-existing cartilage. A majority of the skeleton is formed by endochondral ossification. In the bone formation, three different types of cells are involved: Osteoblasts, Osteocytes and Osteoclasts.
The bone cells, Osteoblasts (immature, bone-forming cells) and Osteocytes (mature bone cells), are derived from osteochondro progenitor cells arose from mesenchymal stem cells (MSCs). In studying these cells, two twist basic helix-loop-helix transcription factor 2 (Twist2 also known as Dermo1) [7172] and paired related homeobox1, rat (Prrx1 also known as Prx1) [73]-containing promoters are used in osteochondro progenitor cells and mesenchymal cells. Also, for the bone growth and physiological studies, bone gamma-carboxyglutamate protein (BGLAP; OC, also known as HOC, OCN, Osteocalcin) [74], collagen, type I, alpha 1 (Col1a1) [75] and Sp7 transcription factor 7 (Sp7, also known as osterix, Osx1) [76] -promoters are used for osteoblasts and Dmp1 (dentin matrix protein 1) is used for osteocytes [77]. The osteoclasts, which is the bone-resorbing cells, are derived from hematopoietic progenitors. For studying the functional regulation of osteoclasts, cathepsin K (Ctsk) promoter is used [75787980].
3) Muscle-specific promoters
The muscular system includes three different types of muscle tissue: skeletal (also called striated), smooth (non-striated) and cardiac muscles. Actin, alpha 1, skeletal muscle (ACTA1 also known as HAS) [84] expressed in striated muscles, myogenic factor 5 (Myf5) [85] and myogenic factor 6 (Myh6 also known as αMHC) expressed in differentiated myocytes [86] are used in studies of skeletal muscle. Creatine kinase, muscle (Ckmm also known as Mck) is expressed in both skeletal and cardiac muscles [87]. Transgelin (Tagln also known as SM22α; smooth muscle protein 22-alpha) is expressed in vascular smooth muscle cells of liver and lung and used in studies of vascular diseases [88].
Digestive system
This system includes digestive tract and various accessory organs to assist digestion by secreting enzymes. The digestive tract (also called gastrointestinal tract) consists of the mouth, esophagus, stomach, small intestine, large intestine and anus. The accessory organs consist of salivary glands, liver, pancreas and so on.
1) Esophagus, Stomach and Intestine-specific promoters
Expression of the ED-L2 promoter of the Epstein-Barr virus, (ED-L2) is reported in squamous epithelial of the tongue, esophagus and forestomach in previous studies [899091]. To study the gastric cancer and diseases of the stomach, the ATPase, H+/K+ exchanging, beta polypeptide (Atp4b) [92], H+,K+-ATPase b-subunit (HK) [93] and trefoil factor 1 (Tff1, also known as PS2 or Bcei) promoters have been used. Atp4b and HK are expressed in progenitors of parietal cells and parietal cells secreting hydrochloric acid (HCl) and intrinsic factor, respectively. Tff1 is expressed in the epithelium of the glandular stomach, a source of secreting gastric enzymes and acid for digestion [94].
The carbonic anhydrase 1 (Car1) is expressed in the large intestinal tissues, including the cecum and colon [95]. This promoter is useful in colon disease research. Villin 1 (Vil1) promoter is most widely used in intestine studies and is expressed in epithelial cells of the small and large intestines from embryonic day 12.5 to adulthood [96].
2) Salivary gland, Liver and pancreas-specific promoters
Achaete-scute family bHLH transcription factor 3 (Ascl3 also known as Sgn1) promoter is expressed in progenitors of both secretory acinar and ductal cells, and thus it is used in developmental studies of salivary gland [97]. Albumin (Alb) promoter is known to exclusively express the Cre recombinase in the liver and are the most commonly used in liver-specific genetic studies [9899].
Pancreatic cells are divided two compartments: alpha, beta, delta, epsilon and PP (pancreatic polypeptide) cells are involved in endocrine function and acinal cells and epithelial cells lining ducts are involved in exocrine function. So far, various pancreatic cell-specific promoters have been identified and used. Glucagon (Gcg, also known as Glu) [100101], ghrelin (Ghrl) [102] and pancreatic polypeptide (Ppy, also known as PP) [101] are used for alpha, epsilon and PP cells, respectively. Insulin I (Ins1, also known as MIP) [103] and insulin 2 (Ins2, also known as RIP) [104] are available in beta cell specific studies. Pancreas specific transcription factor, 1a (Ptf1a) [105] and SRY (sex determining region Y)-box 9 (SOX9) [106] are useful in pancreatic acinal and ductal cell studies, respectively.
Urogenital system
The Urogenital system is composed of urinary system including bladder and kidney and reproductive organs including ovary and testis.
1) Bladder and Kidney-specific promoters
Uroplakin 2 (Upk2) is expressed in urothelium of the bladder and used for the bladder cancer and urothelium functional studies [107]. The promoters for the kidney study are as follows. Aquaporin 2 (Aqp2) is expressed in collecting duct of the kidney and used in nephrological studies [108]. Forkhead box D1 (Foxd1) is detected in metanephric mesenchyme and is useful in studies of pericyte differentiation and fibrotic disease of kidney [109]. The expression of glial cell line derived neurotrophic factor (Gdnf) is restricted in renal progenitor cells and this promoter is useful in kidney developmental studies [110]. Gamma-glutamyltransferase (Ggt1) is expressed in the cortical tubular epithelium of the kidney and used for studying polycystic kidney disease and renal fibrosis [111]. Kidney androgen regulated protein (Kap) is expressed in renal proximal tubule of male kidney [112]. Nephrosis 2 (Nphs2, also known as podocin) is exclusively expressed in podocytes within the kidney glomeruli [113].
2) Ovary and Testis-specific promoters
Growth differentiation factor 9 (Gdf9) is expressed in oocytes of the primordial follicles and used in folliculogenesis and oocyte developmental studies [114]. Expression of the zona pellucida glycoprotein 3 (Zp3) is detected in female germ line, and thus this promoter is useful for generating the deletion of the desired gene in the germ line [114115116]. Expression of the anti-Mullerian hormone (Amh) [117] is restricted in Sertoli cells and used in studies of the male embryonic sexual differentiation and spermatogenesis. Heat shock protein 2 (Hspa2) is expressed in spermatocytes and spermatids within the testis and this promoter is useful in assessing the gene functional studies during and after meiotic prophase in pachytene spermatocytes [118].
Accessing the useful resources of Cre driver lines: Cre portal sites
To date, a huge numerous Cre transgenic mouse lines have been established and being generated continuously. Here we introduce a free, publicly accessible web site that can be used to provide more accurate and versatile information on the Cre mouse lines required by the researchers in Table 3.
Unfortunately, there is still no worldwide database site where researchers can find all the information about the desired Cre mouse lines in one place. The scientific community makes a constant effort for efficiency in collecting and sharing this information. Coordination of resources for conditional expression of mutated mouse alleles (CREATE) consortium web site (http://creline.org/) had integrated and provided information on Cre mouse lines from four international partner databases: Cre portal at MGI (http://www.creportal.org), Cre-X-Mice (http://nagy.mshri.on.ca/cre_new/index.php), CreZoo (http://bioit.fleming.gr/crezoo) and CreERT2Zoo (http://www.ics-mci.fr/crezoo.html) [119120121122123124125126]. But now information of CREATE has integrated into Cre portal at International Mouse Phenotyping Consortium (IMPC) (https://www.mousephenotype.org/data/order/creline). Cre portal sites of MGI and CreZOO are available, but Cre-X-Mice and CreERT2Zoo has integrated into Mouse Resource Browse (MRB) (http://bioit.fleming.gr/mrb) and MouseCre (Mouse Cre and CreERT2 zoo) (http://mousecre.phenomin.fr/), respectively. The Jackson Laboratory's Cre Repository (http://www.Jax.org/research-and-faculty/resources/cre-repository), Neuroscience Blueprint Cre Driver Network of NIH (National Institutes of Health) (http://www.credrivermice.org/) and Gene Expression Nervous System Atlas (GENSAT) (http://www.gensat.org/cre.jsp) can also be used to find information about desired Cre mouse lines. In particular, GENSAT [21127] and Neuroscience Blueprint Cre Driver Network [29] provide useful information in neuroscience.
Conclusions
Technology using Cre-loxp system provides the sophistication to study the gene functions. To date, a tremendous amount of Cre-driver lines has been developed by researchers and are being shared through international networks. Recently, Cre-loxp system for more precise control has been continuously developed. Various types of Cre plasmids are being developed such as fluorescent Cre fused to fluorescent reporter, optimized Cre with high expression ratio and split Cre having the different promoters in N- and C-terminal regions. In addition, Cre-loxp system approaches has been continually being developed using Crispr/Cas9 technology and viral system. Using more sophisticated control techniques, researchers will be able to understand more precise gene functions by studying the function of specific genes at desired time (temporal) and tissue (spatial). The useful Cre-related portal sites and databases will enhance the efficiency of research by allowing researchers to find and obtain the suitable Cre-driver lines for research.



 <0.15,
<0.15,  0.15~2.0,
0.15~2.0,  2.0~4.0,
2.0~4.0,  4.0~6.0,
4.0~6.0,  6.0~8.0 and
6.0~8.0 and  8.0<
8.0<
 XML Download
XML Download