

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Acinetobacter baumannii is a rod-shaped, non-motile gram-negative bacterium. Whereas other Acinetobacter spp. are naturally isolated in soil, water and animal habitats, A. baumannii is intensely isolated in the hospital environment [12]. It causes opportunistic infections in patients with underlying diseases and immunosuppression, which leads to various diseases, such as pneumonia, bacteremia, endocarditis, skin and soft-tissue infections, urinary-tract infection, and meningitis [3]. This bacterium is also referred to as the ‘Iraq bacter’ because it has exploded in US military hospital patients during the Iraq war [4].
About 30 years ago, A. baumannii infections were treated with traditional antibiotics [5]. However, it is now resistant to almost all major antibiotics, including penicillins, carbapenems, and cephalosporins [5]. The increase of these multidrug-resistant (MDR) bacteria is becoming a serious clinical problem, and the incidence is rapidly increasing worldwide. A. baumannii is listed on the World Health Organization's catalog of bacteria for which new treatments are critically needed [6]. Despite its clinical importance, there has been little progress in developing a control strategy for this new threat.
Nucleotide-binding oligomerization domain (Nod)-like receptors (NLRs) are a family of cytosolic receptors that can recognize microbial or damage-associated molecules [7]. Among them, Nod1 and Nod2 are the first identified NLRs and recognize bacterial peptidoglycan derivatives, meso-diaminophimelic acid (DAP) and muramyl dipeptide (MDP), respectively [8]. It has been known that Nod1 is ubiquitously expressed, whereas Nod2 is mostly in immune cells [9]. After recognition, they recruit and associate with an adaptor protein receptor interacting serine/threonine-protein kinase 2 (Ripk2), which activates downstream signaling, such as nuclear factor kappa B (NF-κB) and mitogen-activated protein kinases (MAPKs) [7910]. These events are finally involved in host defense against microbial infections by producing cytokines or inflammatory mediators, such as reactive oxygen species (ROS) [1112].
It has been widely known that Nod1 and Nod2 regulate innate immune response and protect hosts against pulmonary bacterial infections [11131415]. However, little is known about the role of the Nod1 and Nod2 pathways in host immune response against A. baumannii infection. A previous study revealed that the Nod1/2-Ripk2 pathways control intracellular growth of A. baumannii in a human lung epithelial-cell line, A549 cells [16]. The signaling is also essential for NF-κB activation and β-defensin expression in the cells in response to A. baumannii [16]. An in vivo study showed that Nod2 contributes to bacterial clearance and ROS production in the lungs of mice infected with A. baumannii at an early phase of infection [17]. However, we still do not know the function of Nod1 in control of A. baumannii infection in vivo. In this study, we investigated the in vivo role of Nod1 in control of bacterial growth and immune responses in the lungs of mice infected with A. baumannii.
Materials and Methods
Mice
WT C57BL/6 mice were obtained from Koatech (Pyeongtaek, Korea). Nod1−/− mice were gifts from Prof. Gabriel Núñez (University of Michigan, USA) and have been previously described [18]. Protocols for animal studies were approved by the Institutional Animals Care and Use Committee of Chonnam National University (Approval No: CNU IACUC-YB-R-2017-76).
Bacterial preparation
A. baumannii strain KCCM 35453 (ATCC 15150) was purchased from the Korean Culture Center of Microorganisms (Seoul, Korea). Single colonies were inoculated into 10 mL of Luria-Bertani (LB) broth supplemented with ampicillin (50 mg/mL) and grown overnight at 37℃ with 200 rpm shaking. A 1:5 dilution of the culture suspension was allowed to grow in fresh medium at 37℃ with 200 rpm shaking for an additional 2 h. Bacteria were washed and resuspended with sterile phosphate buffered saline (PBS) to a final concentration of 109 colony-forming units (CFU)/mL. Bacteria were diluted to desired concentrations for use in experiments.
In vivo experiments
Mice were anesthetized by intraperitoneal injection of 10 mg/kg Rompun (Bayer, Seoul, Korea) and 50 mg/kg Zoletil (Virbac, Seoul, Korea). They were then intranasally (i.n) inoculated with 30 µL of A. baumannii (1×109 CFU/mL) suspension in PBS. Lung homogenate was collected at 1 and 3 days post infection to quantify bacterial loads, cytokines production, and mouse β-defensins (mBD) expression.
Bacterial counts in lung homogenate
Lung homogenates were spread onto LB agar plates supplemented with ampicillin (50 mg/mL). Following overnight culture at 37℃ in an incubator, bacterial colonies were counted, and the number of bacteria was expressed as CFU/g of lung tissue.
Measurement of cytokines
Concentrations of IL-6, TNF-α, and IL-1β from the lung homogenates of A. baumannii-infected mice or culture supernatant of macrophages were measured using ELISA kits (R&D System, Minneapolis, MN, USA) according to the manufacturer's instructions.
Histopathologic examination
The left lobe of the lung was harvested and fixed in 10% neutral formalin for histopathological observation. The tissues were routinely processed with alcohol and xylene series and embedded in paraffin. Three-micrometer sections were prepared, stained with hematoxylin-eosin (HE), and examined by microscopy. Histopathology of the lung was blindly evaluated using an arbitrary scoring system according to described in a previous study [19].
Quantitative real-time PCR
Total RNA was extracted from lung tissue using easy-BLUE (Intron Biotechnology, Korea) according to the manufacturer's instructions. One microgram of total RNA was reverse transcribed to cDNA by using ReverTra Ace qPCR RT Master Mix and cDNA Synthesis kit (Toyobo, Osaka, Japan). Equal amounts (1 µL) of cDNA were used for real-time PCR on a Rotor-Gene Q (Qiagen, Hilden, Germany) using SYBR Green PCR kit (Qiagen). GAPDH was used for normalization. The following primers were used for real time PCR: GAPDH forward: 5′-CAGTGGATGCAGGGATGATGTTCT-3′; GAPDH reverse: 5′-GTGGAGATTGTTGCCATCAACG-3′; mBD-1 forward: 5′-CCAGATGGAGCCAGGTGTTG-3′; mBD-1 reverse: 5′-AGCTGGAGCGGAGACAGAATCC-3′; mBD-2 forward: 5′-AAGTATTGGATACGAAGCAG-3′; mBD-2 reverse: 5′-TGGCAGAAGGAGGACAAATG; mBD-3 forward: 5′-GCATTGGCAACACTCGTCAGA-3′; mBD-3 reverse: 5′-CGGGATCTTGGTCTTCTCTA-3′; mBD-4 forward: 5′-GCAGCCTTACCCAAATTATC-3′; mBD-4 reverse: 5′-ACAATTGCCAATCTGTCGAA-3′.
Macrophage culture and stimulation
Bone marrow-derived macrophages (BMDMs) were prepared as previously described [20]. The cells were seeded in 48-well plates at the concentration of 2×105/well and incubated in a 5% CO2 incubator at 37℃ overnight. Subsequently, cells were either infected or not infected with A. baumannii at the indicated multiplicity of infection (MOI) by exposure for 60 min, and extracellular bacterial growth was inhibited by gentamicin (50 µg/mL) treatment. Culture supernatant was collected 24 h after infection for cytokine measurement.
Statistical analysis
Difference between groups was assessed by two-tailed Student's t-test or one-way analysis of variance followed by post hoc analysis (Newman-Keuls multiple comparison test). All statistical analyses were performed using GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). Statistical significance was considered at P<0.05.
Results
Bacterial clearance in the lungs of A. baumannii-infected mice
WT and Nod1-deficient mice were intranasally infected with A. baumannii and the bacterial CFUs were measured in the lungs at 1 and 3 days postinfection (dpi). The experiment was repeated twice independently. The bacterial CFUs in the lungs were over 8 log10CFU/g lung at 1 dpi and decreased by 4 log10CFU/g lung at 3 dpi in both WT and Nod1-deficient mice (Figures 1A, B). There was no significant difference of the bacterial CFUs in the lungs between WT and Nod1-deficient mice (Figures 1A, B).
Cytokines production in the lung homogenates of A. baumannii-infected mice
The level of cytokines in the lung homogenates was measured by ELISA. IL-6 and TNF-α levels in WT and Nod1-deficient mice infected with A. baumannii were reduced more on 3 dpi than on day 1, whereas IL-1β production was increased at 3 dpi (Figures 2A–C). Except for TNF-α at day 3, the level of which was higher in the lung homogenate of Nod1-deficient mice, the levesl of each cytokine were comparable between WT and Nod1-deficient mice at 1 or 3 dpi (Figures 2A–C).
Lung pathology in A. baumannii-infected mice
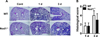
We next examined the role of Nod1 in A. baumannii-induced lung pathology in mice. Intranasal infection of A. baumannii led to moderate to severe lung pathology, which was characterized by neutrophilic infiltration and edematous lesion (Figure 3A). The lung pathology was getting severe at day 3 compared to day 1 in both WT and Nod1-deficient mice (Figures 3A, B). When quantitatively evaluated, there was no significant difference of histological score in lung pathology between WT and Nod1-deficient mice (Figure 3B).
Expression of β-defensins in the lungs of mice infected with A. baumannii
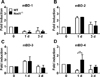
A. baumannii induce gene expression of human β-defensin 2 (hBD-2) in human airway epithelial cells, which is regulated by Nod1 and Nod2 signaling [1621]. Accordingly, we tried to find out whether Nod1 deficiency leads to impaired expression of β-defensins in vivo. mBD-1 expression in the lungs was reduced at days 1 and 3 after A. baumannii infection, compared to that in uninfected control animals (Figure 4A). In contrast, the gene expression of mBD-2 and mBD-3 was increased at day 1 and restored at day 3 (Figures 4B, C). There was no significant difference of gene expression of mBD-1, mBD-2, and mBD-3 in the lungs of WT and Nod1-deficient mice (Figures 4A–C). mBD-4 expression, a mouse orthologue of hBD-2, was also decreased in WT mice at days 1 and 3 compared to the uninfected controls, whereas it did not change in Nod1-deficient mice (Figure 4D).
In vitro cytokine production in A. baumannii-infected macrophages
Macrophages play a role in early host resistance to pulmonary infection by A. baumannii [22], and Nod1 is also functional in macrophages [23]. We then investigated the role of Nod1 in the production of cytokines in response to A. baumannii in macrophages. A. baumannii induced the production of IL-6 and TNF-α in both WT and Nod1-deficient BMDMs without any difference in their levels between the two groups (Figures 5A–C).
Discussion
The role of Nod2 in host defenses against respiratory bacterial infections has been widely studied, whereas studies about Nod1 are limited. Nod2 is knownt to contribute to host resistance against various respiratory pathogens, such as Staphylococcus aureus, Mycobacterium tuberculosis, M. abscessus, Streptococcus pneumoniae, and A. baumannii [131724252627]. Nod2 also has antiviral activity by regulating type I interferons production in response to ssRNA virus via a mitochondrial antiviral-signaling protein (MAVS)-dependent pathway [28]. In Legionella pneumophila-infected mice, Nod1 deficiency led to impaired neutrophilic recruitment to the lungs [1529]. In the study, a single deficiency of Nod1 or Nod2 did not influence the clearing of L. pneumophila from the lungs of mice at most time points [1529], except on day 3, at which the bacterial CFUs in the lungs were higher in Nod1-deficient mice than in WT mice [15]. However, the bacterial clearing was impaired in mice deficient in Ripk2 [29], which is an essential adaptor protein of both Nod1 and Nod2, indicating that Nod1 and Nod2 cooperate to control the growth of L. pneumophila in the lungs.
In our study, we have shown that Nod1 is not required to clear bacteria from the lungs of mice infected with A. baumannii. Nod1 deficiency did not affect in vivo cytokines production, lung pathology, or β-defensins expression in the lungs of infected mice. Moreover, A. baumannii-induced production of IL-6 and TNF-α was not impaired in Nod1-deficient macrophages. These results are similar to those in a study of Pseudomonas aeruginosa infection [30]. When infected with P. aeruginosa intranasally, neutrophils recruitment and production of chemokines, such as CXCL1 and CXCL2, in broncholavage (BAL) fluids were comparable between WT and Nod1-deficient mice [30]. Moreover, Nod1 is not involved in bacterial clearing in BAL fluids of mice infected with P. aeruginosa [30]. Taken toghether, Nod1 seems to not have a significant effect on host defense against acute respiratory infection of gram-negative bacteria with strong TLR4 activity.
There are some considerations about the possibility that Nod1 might not play an important role in host defense against pulmonary infection of A. baumannii. Although epithelial Nod1 is involved in β-defensin expression in response to the bacteria in vitro condition [16], our results showed that Nod1 is not needed for mBD-2 and mBD-3 expression in the lungs of infected mice. In addition, hBD-2 in human breast milk inhibits some bacterial growth, including Serratia marcescen and P. aeruginosa, with minimum inhibitory concentrations (MICs) of <0.5 mg/mL, whereas it showed a higher MIC (4 mg/mL) against a multidrug-resistant strain of A. baumannii [31], indicating that β-defensins may not be able to control in vivo growth of A. baumannii. Therefore, the effect of Nod1 on in vivo expression of β-defensins against A. baumannii is likely very limited, and at most, modest in the bacterial growth control by β-defensins.
We should also consider the role of Nod1 in the function of macrophages and neutrophils. At 4 h after A. baumannii infection, macrophages are dominant in BAL fluids, whereas neutrophils are at 24 h after infection [22]. A Nod1 ligand iE-DAP can induce the production of IL-6 and TNF-α in murine macrophages, but very low levels (<60 pg/mL) compared to those by LPS [23]. For bacteria possessing strong TLRs-stimulatory activity, it is likely that TLR2 or TLR4 masks the role of Nod1 and Nod2 in immune responses in macrophages. In fact, Nod1, Nod2, and Ripk2 are more critical for cytokines production in macrophages in response to several gram-negative bacteria when there is TLRs tolerization or deficiency [323334]. In this study, Nod1 deficiency does not influence cytokines production in A. baumannii-infected macrophages. Our previous study showed that A. baumannii-induced production of IL-6 and TNF-α was mostly abolished in TLR4-deficient macrophages [35]. Therefore, Nod1 is likely irrelevant in the immune response of macrophages against A. baumannii in the presence of TLR4.
There is a controversy about the effect of Nod1 in regulating neutrophil function. Clarke et al. showed that recognition of microbiota peptidoglycan by Nod1 strengthens neutrophil ability to kill some bacteria, such as S. pneumoniae and S. aureus [36]. However, Nod1 is not expressed in either human or mouse neutrophils, whereas a strong expression of Nod2 is observed [3738]. Nod2 agonist MDP induces cytokines and chemokines in neutrophils, but Nod1 stimulation does not [3738]. Further studies seems to be necessary to clarify the role of Nod1 in host defense by neutrophils against bacterial infection.
In conclusion, our study revealed that a single deficiency of Nod1 does not affect in vivo host defense against pulmonary infection of A. baumannii, although it is involved in NF-κB activation, hBD-2 expression, and restriction of the intracellular bacterial growth in respiratory epithelial cells against A. baumannii infection [16]. It cannot be excluded that Nod1 cooperates with Nod2 or TLRs to regulate host immune response against A. baumannii infection.

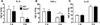

 XML Download
XML Download