

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Cancer, obesity, and innate immune deficiency are major diseases whose incidence has rapidly increased in recent years, leading to risings costs of treatment and deterioration of quality of life for those affected [1037]. One protein that has been implicated to play a role in all these conditions is PLAC8, a 12.5-kDa protein consisting of 112 amino acids, which is encoded by the Plac8 gene and expressed primarily in epithelial and immune cells [922]. Specifically, a number of previous studies conducted using embryonic stem (ES) cell-based Plac8 knockout (KO) mouse models have reported the involvement of PLAC8 in colorectal and pancreatic cancer progression [161924], irregular differentiation of brown/white fat cells [1315], and innate immune deficiency due to various mechanisms [22].
Colorectal and pancreatic cancers are devastating diseases that show poor survival rates due to a low incidence of early stage detection, rapid metastasis, and poor response to therapeutics [2627]. Numerous genetic studies have reported frequent mutations in the KRAS and TP53 genes in colorectal and pancreatic cancers, which promote strong invasiveness via the epithelial to mesenchymal transition (EMT) [231725]. Notably, it was found that PLAC8 accelerates tumorigenesis and metastasis by promoting cell growth in the RAS and TP53 mutant types of pancreatic and colorectal cancers [1619]. In addition, invasiveness and tumor size are increased by PLAC8 upregulation through the activation of extracellular signal-regulated kinase 2 (ERK2) in colorectal cancer [24]. Obesity, which afflicts approximately 13% of individuals worldwide [20], is characterized by excessive fat tissue that is associated with an increase in both adipocyte number and size [7]. PLAC8 has further been shown to modulate the morphology and thermogenic ability of brown adipocytes by acting as a novel upstream regulator of CCAAT/Enhancer binding protein β (C/EBPβ). Loss of PLAC8 also increases the size of white adipocytes through the induction of the early growth response 2 (EGR2/KROX20) and kinesin family member 4 (KIF4) proteins, which are the early regulators of white adipogenesis [1315]. The innate immune response is a major host defense system that recognizes, attacks, and eliminates foreign and infectious substances from the body [21]. Neutrophils, the most abundant cell type in the innate immunity, recognize various pathogens and control immune responses, mainly by chemotaxis and phagocytosis [6113336]. PLAC8 has been implicated in neutrophil function, and a Plac8 KO impairs host defense activity by accelerating apoptosis and reducing phagocytic ability of neutrophils [22]. Collectively, these findings highlight the potential utility of an unmarked Plac8 KO mouse model for the investigation of tumorigenesis, adipogenesis, and innate immunity.
Engineered nucleases have been utilized to generate a number of rapidly developing tools for genome editing [4]. A system of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated (Cas) proteins is the third-generation of an engineered endonuclease, containing a guide RNA and Cas protein, which efficiently introduces DNA double-strand breaks at a target site, leading to non-homologous end joining (NHEJ)-mediated DNA repair [38]. Compared with conventional gene targeting strategies in ES cells, methods using CRISPR/Cas systems enable the manipulation of genes at the single nucleotide level and show increased efficiency for establishing animal models, mimicking human disease phenotypes, in a short period of time [29]. Here, to avoid any conflicts caused by patents or primary authority, we generated a new Plac8 KO mouse model that does not harbor a negative selection marker, such as the neomycin resistance (NeoR), using the CRISPR/Cas9 system. We expect that this model will help to further elucidate the myriad of roles that PLAC8 plays in human disease.
Materials and Methods
In vitro synthesis of single guide RNAs and Cas9 mRNA
Cas9 mRNA and single guide RNAs (sgRNAs) were synthesized as described previously [534]. Briefly, Cas9 mRNA was prepared in vitro from linear DNA templates, using the mMESSAGE mMACHINE T7 Ultra Transcription Kit (Invitrogen, Carlsbad, CA, USA), and sgRNAs were generated using the MEGAshortscript T7 Transcription Kit (Invitrogen), both according to the manufacturer's instructions. The following sequences were used for sgRNA synthesis: left sgRNA, TTGAGACCTCGCATCGAAGATGG; right sgRNA, TGTGATTGCTTCAGTGACTGCGG.
Generation and maintenance of Plac8 KO mice
Plac8 KO mice were generated using the CRISPR/Cas9 system, as described previously [32]. In brief, Cas9 mRNA and sgRNAs were microinjected into fertilized embryos of C57BL/6J mice. Mutations in Plac8 were confirmed by Sanger sequencing analyses (BIONICS Ltd., Korea). Homozygous KO mice were born from a heterozygous intercross and used for phenotypic analyses in parallel with age- and sex-matched wild-type (WT) littermates as a control group. All mice were genotyped 2 weeks after birth, using PCR with specific primers (forward, 5′-GCAACTTTCCACACAGTAAG-3′ and reverse, 5′-GCACAGCAGGTAGGGCTAAC-3′), and animals were housed according to their gender after weaning. Mice were kept in an enriched environment under the standard conditions (22±2℃ temperature, 40–60% humidity) with a 12 h light/dark cycle at the Specific-Pathogen-Free (SPF) facility [18]. During the experiment period, mice were given free access to food (Lab-Diet 5002 Certified Rodent Diet, PMI Nutrition International, St. Louis, MO, USA) and water. All experiments in this study were approved by the Institutional Animal Care and Use Committees at Yonsei University and Seoul National University Hospital. Animals were maintained in accordance with Guide for the Care and Use of Laboratory Animals, 8th edition, National Research Council (2010).
RNA isolation and Real-time quantitative PCR (RT-qPCR)
Total RNA was extracted from the intestine, liver, heart, lung, and brown adipose tissues (BAT) of each mouse using TRIzol reagent (Invitrogen) as described previously [31]. Subsequently, cDNA was synthesized using a RevertAid First strand cDNA Synthesis Kit (Thermo Fisher Scientific). Real-time quantitative PCR was performed in the triplicate using CFX (Bio-Rad) [30] with following primers; Plac8, 5′-GTCTTGGATGTCAAGTGGCAGC-3′ (forward) and 5′-CAGGACAGAAGAGTGTGACCATG-3′ (reverse); and Gapdh, 5′-CATCACTGCCACCCAGAAGACTG-3′ (forward) and 5′-ATGCCAGTGAGCTTCCCGTTCAG-3′ (reverse).
Measurement of physiological parameters: body weight gain, food, and water consumption
The main physiological parameters of Plac8 KO mice and their WT littermates were monitored from 4 to 52 weeks of age. During that time period, body weight and both food and water consumption were measured once a week, with daily observation of clinical signs. Food and water consumption were calculated as the amount of intake per mouse for 24 h. All experiments were conducted during the light cycle with gentle handling, so as to minimize stress on the animals. The raw data are available on request.
Hematology, serum biochemistry, urinalysis, and organ weight measurement
All mice were sacrificed by anesthetizing with isoflurane and exsanguination at 52 weeks of age. Whole blood was collected from the inferior vena cava in K2EDTA-containing tubes (BD Microtainer, Becton Dickinson, Franklin Lakes, NJ, USA) and analyzed using the ADVIA 2120i Hematology System (SIEMENS Healthineers, Erlangen, Germany) for determination of the following parameters: total and differential white blood cell, red blood cell (RBC), platelet (PLT), hemoglobin (HGB), and hematocrit (HCT) counts, as well as mean corpuscular volume, mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC). In serum, the levels of blood urea nitrogen, total cholesterol, total protein, total bilirubin, aspartate transaminase (AST), alkaline phosphatase (ALP), alanine transaminase (ALT), γ-GT, albumin, creatinine, triglyceride (TG), glucose, albumin/globulin (A/G) ratio, potassium, chloride, sodium, and phosphorus were measured with a Hitachi 7070 Chemistry Analyzer (Hitachi, Japan). Urine collected from each mouse was applied to a urine stick (Multistick 10 SG, SIEMENS) and analyzed using an automatic urinalysis machine (CLINITEK Advantus Urine Chemistry Analyzer, SIEMENS) for measurement of leukocytes, nitrite, urobilinogen, protein, pH, blood, specific gravity, ketones, bilirubin, and glucose. The weight and gross morphological changes of all organs were examined after exsanguination. Testis and epididymis were preserved in Bouin's solution, and Harderian glands and eyes were fixed in Davidson solution. All other organs were fixed in 10% neutral buffered formalin.
Statistical analysis
All values are expressed as the mean±standard deviation (SD). Statistical analysis was carried out using Welch's t-test in SPSS software, version 22.0 (SPSS Inc., Chicago, IL, USA). P-values <0.05 were regarded as statistically significant.
Go to : 
Results
Establishment of a Plac8 KO mouse model by CRISPR/Cas9 microinjection
To construct a Plac8 KO mouse model, we designed sgRNAs targeting the translation start codon in exon 2 of the Plac8 gene [812]. These were delivered to mouse embryos by microinjection together with Cas9 mRNA, and Plac8 gene mutations were screened by PCR, using primers spanning the target sites; KOs were confirmed by Sanger sequencing analyses (Figure 1A). The chosen mouse line harbors a 99 bp deletion mutation, abolishing the conventional translational start site (Figure 2A), and genotypes were confirmed by PCR genotyping (Figure 1B). We note that this 99 bp difference was sufficient to distinguish PCR products from WT and KO mice. Analysis of Plac8 transcript level by RT-qPCR showed the impaired Plac8 expression in the intestine, liver, heart, lung, and brown adipose tissues of the Plac8 KO, compared to control mice (Figure 1C). Compared with previously generated Plac8 KO mice, in which exon 4 was replaced with the NeoR cassette, our newly established Plac8 KO mouse model targets an upstream region (exon 2) and contains no selectable marker.
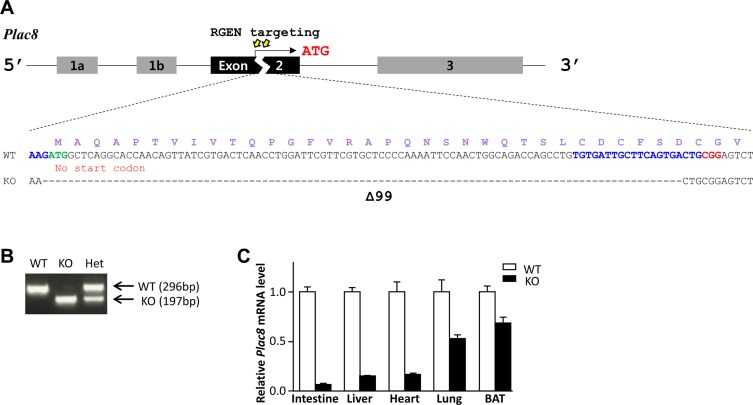 | Figure 1CRISPR/Cas9-mediated generation of a Plac8 KO mouse model. (A) A representative view of the CRISPR/Cas9 targeting strategy used for generating Plac8 knockout (KO) mice. The nucleotide and amino acid (in purple) sequences of wild-type (WT) and KO alleles are shown. Exons are indicated with black and gray boxes, and introns are noted by black lines. The targeting sites are indicated by yellow arrows, and the binding sites for single guide RNAs (sgRNAs) are marked in blue, with the protospacer adjacent motif (PAM) sequence in red. The conventional start codon (ATG) is indicated in green. (B) A representative PCR genotyping result for Plac8 WT, homozygous (KO), and heterozygous (Het) KO mice. (C) Relative Plac8 mRNA levels in the intestine, liver, heart, lung, and brown adipose tissue (BAT) of the Plac8 KO (Black bar) mice were compared to WT (White bar). The mRNA levels were normalized by Gapdh. Graph shows mean±standard deviation.
|
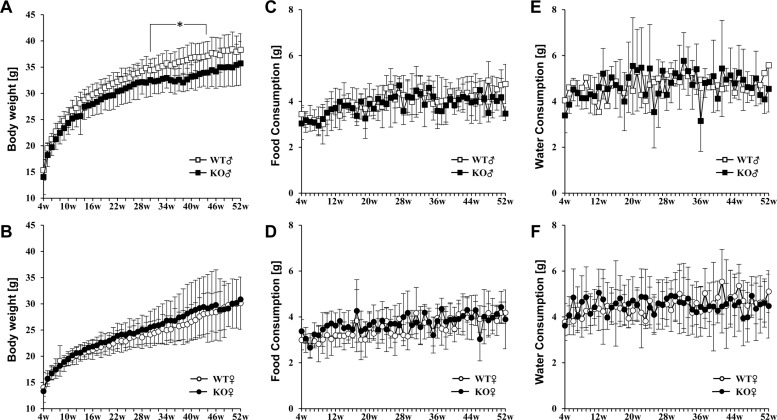 | Figure 2Measurement of physiological parameters in Plac8 KO mice and comparison to WT littermate controls. (A and B) Body weight changes in Plac8 WT and KO of male (A) and female (B) mice from 4 to 52 weeks of age. (C–F) The amount of daily food (C and D) and water (E and F) consumed by WT and Plac8 KO male (C and E) and female (D and F) mice from 4 to 52 weeks of age. WT animals are shown in white dots, and KO mice are shown in black dots. Male mice are indicated with squares (A, C, and E), and female mice are indicated with circles (B, D, and F). Graphs show mean± standard deviation. *P<0.05.
|
Characterization of basic phenotypes in Plac8 KO mice
To characterize the basic phenotypes of Plac8 KO mice, physiological parameters, including genotype and sex ratio at birth, as well as whole body and organ weight gain were examined. The sex ratio of pups born from Plac8 heterozygous knockout (HT) parents was 1:0.9 (male, n=56; female, n=51), whereas the genotype ratio of siblings was 1:2.1:0.6, for WT, HT, and KO, respectively. These data suggest that Plac8 KO does not affect gender determination, although the genotype birth ratio may warrant further investigation. As the previous Plac8 KO mouse exhibited increased adipose tissue weight, we expected the augmentation of body or tissue size and weight in our Plac8 KO mouse model. However, male Plac8 KO mice showed lower body weight compared to their WT littermates over the entire observation period, with significant differences observed between 29 to 44 weeks of age, excepting for week 30 (Figure 2A and Table 1). Conversely, the body weights of female mice were comparable between the two groups (Figure 2A). Because dietary intake is a major factor for body weight gain, food and water consumptions were monitored throughout the experimental time period, and we found no intake differences in Plac8 KO mice, as compared with their WT littermates (Figures 2B and C). Additionally, consistent with previously reported data, no significant changes were observed in any of the other organs (Table 1) [22], although marginal differences were noted in some.
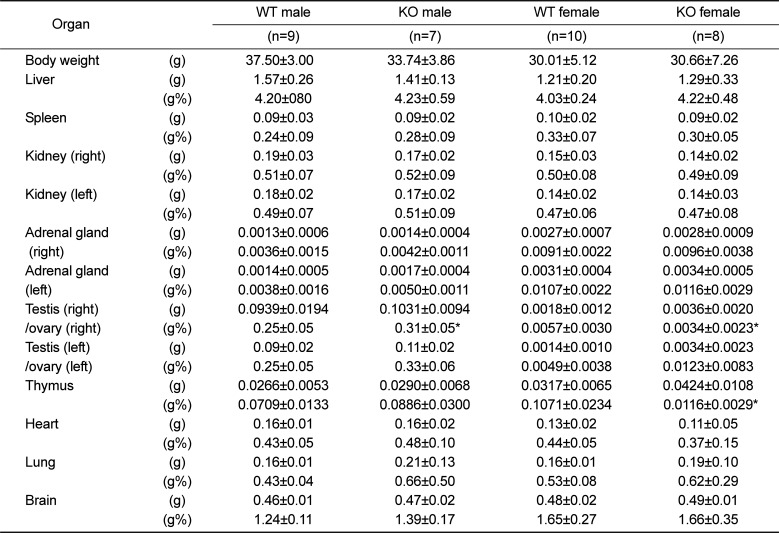
Table 1
Absolute and relative organ weight in 52-week-old Plac8 KO mice
![]()
Hematology, serum biochemistry, and urinalysis in Plac8 KO mice
Analysis of whole blood revealed that RBC, HCT, and PLT counts were increased, whereas MCH and MCHC were decreased in 52-week-old female KO mice, as compared to age and sex-matched WT littermate controls (Table 2). In contrast, all other parameters were comparable between WT and KO mice, except for a decreased population of monocytes in male KO mice vs. controls. Serum biochemistry and urinalysis revealed no changes in Plac8 KO mice compared to their WT littermates (Tables 3 and 4).
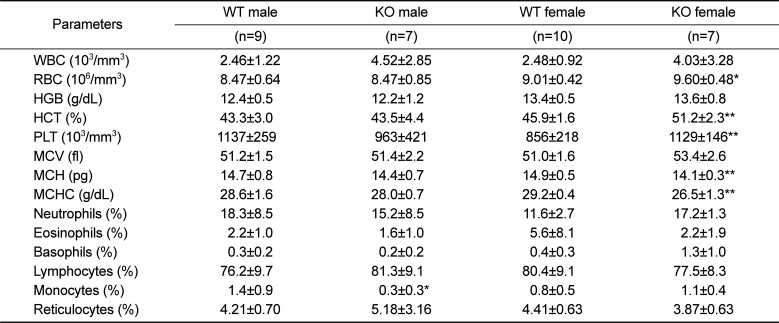
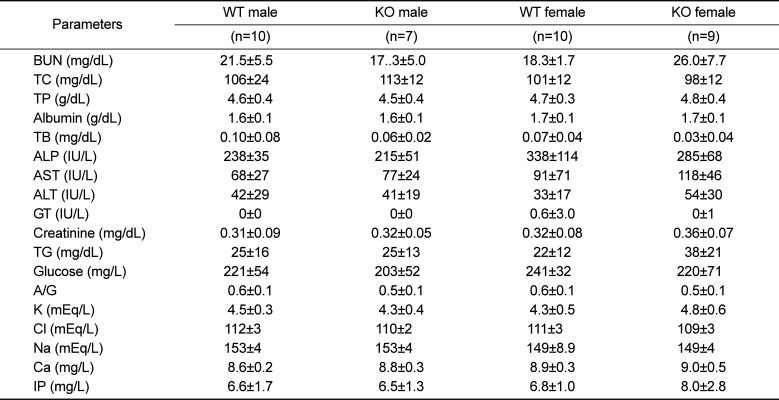
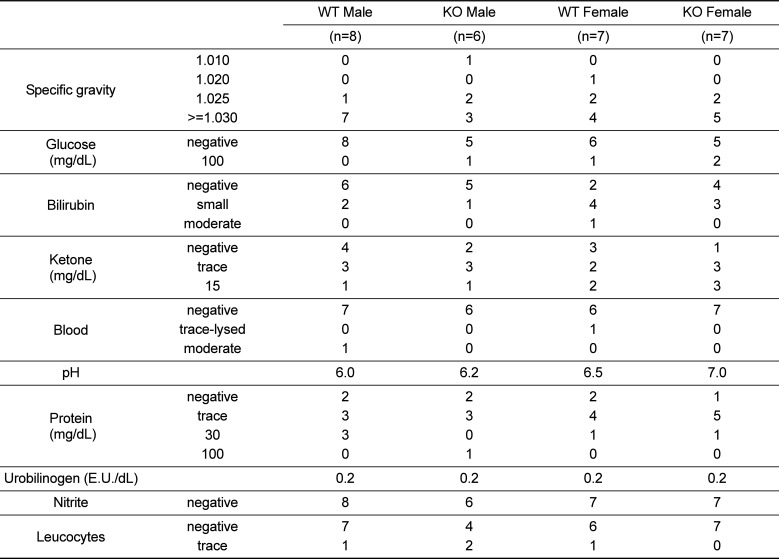
Table 2
Hematological analysis in 52-week-old Plac8 WT and KO mice
![]()
Table 3
Serum biochemistry in 52-week-old Plac8 WT and KO mice
![]()
Table 4
Urinalysis in 52-week-old Plac8 WT and KO mice
![]()
Go to :
Discussion
In this study, we used the CRISPR/Cas9 system to generate an unmarked Plac8 KO mouse line. Macroscopic examinations showed no gross differences in organ size and weight, blood cell counts, serum biochemistry, or urinalysis in KO mice compared to sex-matched WT littermates, although minimal alterations, such as in body weight, were detected in some KO animals under normal conditions. Notably, an observation in the previous Plac8 KO mouse that displays increased weight of whole body and adipose tissues [1314] is an opposite tendency to our Plac8 KO mouse model. It is unclear why there are phenotypic differences between our model and the previously generated model that showed obese phenotype [13], but some possibilities include the distinct targeting loci used in each KO, the presence of the NeoR cassette in the previous model, and mouse background [2839]. Further examinations are required to reveal the phenotypic differences between the previous and our Plac8 KO mouse models. In addition, there is a possibility of unexpected expression of truncated Plac8 protein through an alternative translation by the other in-frame ATG site in exon 3 and 4. As a considerable quantity of Plac8 transcript was detected in the lung and BAT of Plac8 KO mouse, further experiments are required to decide the probable expression of truncated PLAC8 protein in that tissues.
Screening of ES cells with the positive selection reagent, geneticin (G418), is a convenient method for detecting targeted ES cell lines containing the NeoR cassette and has been successfully applied in many instances for KO mouse generation. Despite these results, possible undesirable effects due to the NeoR gene should be considered for improvement of current animal models. For instance, insertion of this gene silences the proximal promoters by five- to ten-fold, regardless of the orientation [1]. Further, NeoR expression can render the cells less glycolytic and change the expressions of genes, such as procollagen 1α, fibronectin, and C-MYC [35]. Thus, our newly established Plac8 KO model may more clearly show the in vivo function of Plac8, free from the off-target effects of the NeoR gene.
We note, however, that in our model, it is unclear whether the lack of a conventional translation start site allows the mutant mRNA to use an alternative ATG codon for translation initiation. However, in this event, the alternative translation would lead to a premature termination codon (PTC) 12 bp downstream of the ATG site, resulting in the probable synthesis of a peptide that is only five amino acids. In contrast, the WT protein is 112 amino acids. We would also expect this truncated mutant transcript to be eliminated by PTC-triggered nonsense-mediated mRNA decay [23].
In conclusion, PLAC8 is a multi-functional protein that is associated with a number of important diseases, such as cancer, obesity, and immune deficiency, that continue to adversely affect large numbers of people but remain incompletely understood [61525]. In addition to the features indicated above, international mouse phenotyping consortium (IMPC) described novel phenotypes of Plac8 KO mouse such as abnormal lens morphology, cataract, abnormal coat appearance, and decrease grip strength (http://www.mousephenotype.org). Based on our and IMPC results, we expect that our unmarked Plac8 KO mouse model, generated using the CRISPR/Cas9 system, will be an important resource for fundamental research into these conditions, as well as for the development of therapeutics. The mouse model is available at the Korea Ministry of Food & Drug Safety (www.nifds.go.kr).
Go to :
 XML Download
XML Download