

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Breast cancer is the second most common cancer in Korean women [1], with 19,219 cases diagnosed in Korea in 2015 [2]; it is also the second leading cause of cancer-related death among women worldwide [3]. Developing and applying mouse models, using both human tumor xenograft models and genetic modifications, have led to our current understanding of molecular mechanisms that operate during breast cancer progression and metastasis [4]. Breast cancers are classified into five intrinsic subtypes based on hormonal and Human Epidermal growth factor Receptor-2 (HER2) receptor status, namely luminal A (Estrogen Receptor (ER)/Progesterone Receptor (PR)-positive major problem), luminal B (ER/PR-positive, HER-negative, high Ki-67, higher histological grade than luminal A), ER-negative/HER2-positive, HER2-positive, and basal-like triple-negative (ER/PR/HER2-negative) [5]. Treatment of human breast cancer patients is based on the hormone receptor status, specifically ER, PR, and HER2 [67], and these hormonal therapies are effective for most patients with hormonal receptor-positive breast cancer; for example, tamoxifen for ER-positive breast cancer and Pertuzumab (Perjeta), Trastuzumab (Herceptin), and Docetaxel (Taxotere) for HER2-positive breast cancer [891011]. However, primary and acquired resistance to hormonal treatments remains to be resolved [101213]. Xenograft models are used to elucidate the underlying mechanisms of resistance, and genetically engineered mouse models (GEMMs) are also useful to understand the mechanisms involved in the pathogenesis and molecular processes of breast cancer and metastasis. In this review, we summarize several mouse models used in breast cancer research and drug development, and their contribution to understanding the molecular pathways in tumorigenesis and metastasis, providing us invaluable insights into possible developments of innovative cancer therapy [14].
Tumor transplantation models
Cell-derived xenografts (CDX), patient-derived xenografts (PDX), and a syngeneic model are well-established tools for evaluating therapeutic efficacy and toxicity and for applying to preclinical assessment (Table 1). CDX transplantation models are useful for investigating breast cancer and metastatic progress [15]. These models are also convenient for investigating genetic alterations that are associated with tumor initiation and growth, but unfortunately show poor clinical predictability.
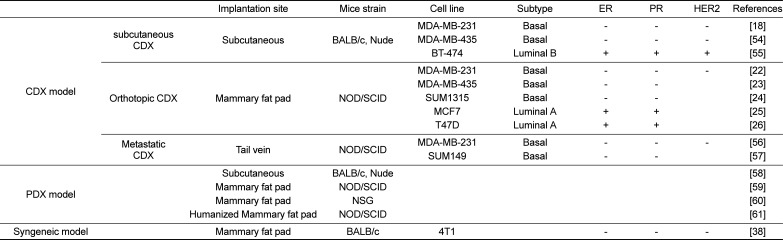
Table 1
Brief overview of breast cancer cell line-derived xenograft (CDX), patient-derived xenograft (PDX) and syngeneic mouse models
| Implantation site | Mice strain | Cell line | Subtype | ER | PR | HER2 | References | ||
|---|---|---|---|---|---|---|---|---|---|
| CDX model | subcutaneous CDX | Subcutaneous | BALB/c, Nude | MDA-MB-231 | Basal | − | − | − | [18] |
| MDA-MB-435 | Basal | − | − | − | [54] | ||||
| BT-474 | Luminal B | + | + | + | [55] | ||||
| Orthotopic CDX | Mammary fat pad | NOD/SCID | MDA-MB-231 | Basal | − | − | − | [22] | |
| MDA-MB-435 | Basal | − | − | [23] | |||||
| SUM1315 | Basal | − | − | [24] | |||||
| MCF7 | Luminal A | + | + | [25] | |||||
| T47D | Luminal A | + | + | [26] | |||||
| Metastatic CDX | Tail vein | NOD/SCID | MDA-MB-231 | Basal | − | − | − | [56] | |
| SUM149 | Basal | − | − | [57] | |||||
| PDX model | Subcutaneous | BALB/c, Nude | [58] | ||||||
| Mammary fat pad | NOD/SCID | [59] | |||||||
| Mammary fat pad | NSG | [60] | |||||||
| Humanized Mammary fat pad | NOD/SCID | [61] | |||||||
| Syngeneic model | Mammary fat pad | BALB/c | 4T1 | - | - | - | [38] | ||
![]()
CDX transplantation models, in which tumor cells are subcutaneously transplanted in nude mice, facilitate straightforward monitoring of tumor growth. Cancer cells in these models show primary tumor growth but do not metastasize [16]. For example, models using BT474 cells (luminal B) [17], as well as MDA-MB-231 [1819] and MDA-MB-435 cells (triple negative) are well-established [20]. Orthotopic CDX transplantation models in which tumor cells are transplanted to a mammary fat pad of NOD/SCID mice are suitable for studying metastatic functions of genes of interest [21] and to investigate malignant and metastatic phenotypes in mice [4]. Several cancer cell lines, such as MDA-MB-231, MDA-MB-435, and SUM1315 (triple negative), as well as MCF7 and T47D cells (luminal A), are used to generate spontaneous metastasis models of breast cancer through orthotopic injection [2223242526]. Metastatic CDX transplantation models in which cancer cells, such as MDA-MB-231 and SUM149 cells, are injected into mouse tail veins are suitable for monitoring experimental metastasis. Taken together, these CDX models allow validation of target genes of interest, and facilitate evaluation of a candidate anti-cancer drug and therapeutics for breast cancer.
In PDX transplantation models, primary human breast carcinomas or tumor fragments are implanted into immune-deficient mice (e.g., nude, NOD/SCID, or NOD/SCID/IL2-receptor null (NSG) mice) subcutaneously or orthotopically [27]. Resulting tumors have characteristics that are similar to those of the original patients' tumors with respect to histology, genomic signature, and heterogeneity, as well as high predictive drug response [142829303132]. These models are used to identify biomarkers for personalized drug selection, and to overcome the limitation of CDX transplantation in clinical therapies [33]. While subcutaneous PDX transplantation models have been used in studies to measure primary tumor growth, orthotopic PDX transplantation models are suitable for mechanistic studies of metastasis and therapeutic resistance [34].
Syngeneic mouse models in which murine cancer cells, such as 4T1, are injected into immune-competent mice (e.g. BALB/c) show more effective metastasis, with characteristics similar to those of breast cancer patients. Advantages of these models over CDX transplantation models include the use of immune-competent mice with normal immune cells and immune system, enabling investigation and development of various immune therapies, for example, with anti-PD-1/PD-L1. These models are also useful to investigate the anti-tumor and anti-metastatic effects of multiple drugs due to the high invasiveness of murine cancer cells [3536373839]. While CDX transplantation models, in which human cancer cells are injected into immunocompromised mice, are well-established in the study of tumor growth and metastasis and the validation related gene profiles, human cancer cells in these models poorly metastasize and show poorly predictable metastatic characteristics.
Collectively, CDX, PDX, and syngeneic models are well-established for evaluating therapeutic efficacy and toxicity and for applying to preclinical assessment. These models could be critical tools for understanding breast cancer progression, and for evaluating responses to targeted therapeutics, in order to predict therapeutic outcomes for breast cancer patient.
Go to : 
Genetically engineered mouse models
GEMMs of breast cancer have greatly contributed to cancer research, improving understanding and validating human cancer genes, genetic pathways, and therapeutic approaches for cancer, as well as investigating cancer progression and metastasis [40]. They are also essential tools for gene expression profiling in breast cancer progression and metastasis. Histopathological features in over 25 different murine GEMMs of breast cancer have been reported [41]. Promoters frequently used in GEMMs of breast cancer are the mouse mammary tumor virus-long terminal repeat (MMTV-LTR), C3(1), and the whey acidic protein (WAP) promoter, which are used to drive mammary expression of oncogenes, such as neu/ErbB2, cyclin D1, Ras, Myc, and Wnt1 [42]. Promoters, origins, activations, active proteins, incidences, latencies, and pathology subtypes of GEMMs of breast cancer are summarized in Table 2.
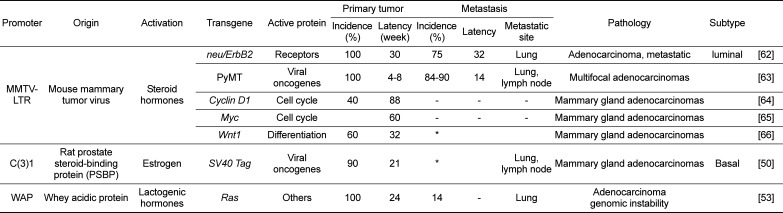
Table 2
Examples of genetically engineered animal tumor models in breast cancer
| Promoter | Origin | Activation | Transgene | Active protein | Primary tumor | Metastasis | Pathology | Subtype | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Incidence (%) | Latency (week) | Incidence (%) | Latency | Metastatic site | ||||||||
| MMTV-LTR | Mouse mammary tumor virus | Steroid hormones | neu/ErbB2 | Receptors | 100 | 30 | 75 | 32 | Lung | Adenocarcinoma, metastatic | luminal | [62] |
| PyMT | Viral oncogenes | 100 | 4-8 | 84-90 | 14 | Lung, lymph node | Multifocal adenocarcinomas | [63] | ||||
| Cyclin D1 | Cell cycle | 40 | 88 | - | - | - | Mammary gland adenocarcinomas | [64] | ||||
| Myc | Cell cycle | 60 | - | - | - | Mammary gland adenocarcinomas | [65] | |||||
| Wnt1 | Differentiation | 60 | 32 | * | Mammary gland adenocarcinomas | [66] | ||||||
| C(3)1 | Rat prostate steroid-binding protein (PSBP) | Estrogen | SV40 Tag | Viral oncogenes | 90 | 21 | * | Lung, lymph node | Mammary gland adenocarcinomas | Basal | [50] | |
| WAP | Whey acidic protein | Lactogenic hormones | Ras | Others | 100 | 24 | 14 | - | Lung | Adenocarcinoma genomic instability | [53] | |
![]()
ErbB2 (neu/HER2) is the EGFR family of receptor tyrosine kinases (RTKs) [43], which form homo- and hetero-dimers in response to ligand stimulation, leading to proliferation, differentiation, cell survival, and apoptosis. ErbB2 functions as a prognostic marker, especially in tumors from patients with lymph node metastasis, and predicts poor survival and early relapse in patients [4445]. Transgenic mice develop multifocal adenocarcinomas by 30 weeks of age, which metastasize to the lungs [4647].
Mammary gland-specific expression of the polyoma middle T antigen (PyMT) -under the control of the MMTV promoter- exhibits multifocal adenocarcinomas with a short median latency, high penetrance, and metastasis to the lungs and lymph nodes. These transgenic mice develop palpable tumors at 4–8 weeks of age, and 84–90% of them show pulmonary metastasis by 14 weeks of age [46]. Pathology of these mice is very similar to that of human breast cancer, regarding hyperplasia, adenoma, and early or late carcinoma [46].
Cyclin D1 has been known to play a critical role in the development of normal alveolar mammary gland tumors induced by c-neu, v-Ha-ras, and other oncogenes. As the expression of Cyclin D1 gene is associated with poor prognosis of estrogen receptor (ER) positive patients, which is supported by relapse-free and overall survival, tumor growth in MMTV-Cyclin D1 models is expected to be ER-positive and estrogen-dependent [4849]. In MMTV-Cyclin D1 mice, mammary adenocarcinoma develops by 22 months of age, which is observed in 40% of the transgenic mice. As Cyclin D1 is a weak oncogene, its expression causes long latency and low incidence of breast cancer in the transgenic mice, and co-expression with potent oncogenes is required for carcinogenesis.
Wnt signaling in human breast cancer has been known to be related to the overexpression of β-catenin. It was also reported that human breast cancers highly express β-catenin which is correlated with poor prognosis of breast cancer patients. In MMTV-Wnt1 transgenic mice, mammary adenocarcinoma develops by 32 weeks of age, which is observed in 60% of the mice, and metastasis to lymph nodes and lungs is observed.
In C3(1)/Tag mice, the SV40 large T-antigen (Tag) is expressed by the regulatory control of the rat prostatic steroid binding protein C3(1) gene, leading to development of prostate tumors in male mice or mammary gland adenocarcinomas in female mice [50]. Characteristics of these models include short latency (~21 weeks) and high penetrance (>90%) (Table 2), which is useful for colony management, as well as nonclinical therapeutic trials. In the transgenic mice, mammary adenocarcinoma develops by 21 weeks of age, which is observed in 90% of the mice. Identification of conserved gene expression features and DNA somatic alterations between the C3(1)/Tag models and human breast tumors suggests that this transgenic model recapitulates human Basal-like cancer (BLBC) [51].
WAP (whey acidic protein) promoter is activated by lactogenic hormones in mammary tumors of mice. Ras oncogene has been known to contribute to human cancer development, and the expression of H-ras oncogene driven by the WAP promoter causes genomic instability, adenocarcinoma, and pulmonary metastasis in mice [5253]. In the transgenic mice, mammary gland tumors develop at 24 weeks of age (Table 2).
Taken together, GEMMs of breast cancer are essential nonclinical models to understand the expression profile of target genes and to validate novel therapeutic strategies.
Go to :
Conclusion
Breast cancer is the most common type of cancer in females. Despite recent advances in its diagnosis and effective therapeutic strategies, further investigations into tumorigenesis, metastasis, and resistance are urgently required. In this review, we provide an overview of tumor transplantation models and GEMMs in order to understand the molecular mechanisms underlying breast cancer progression and metastasis, and to validate the association of human breast cancer with clinical therapeutic trials. Technological advances in order to develop novel mouse models would give us new insights for developing innovative breast cancer therapeutics.
Go to :
 XML Download
XML Download