

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The origin of bile acids is cholesterol in the liver. By numerous cytochrome P450 enzymes, including CYP7A1, CYP8B1 and CYP27A1, bile acids are converted from cholesterol in the liver [1234]. Since bile acids are hydrophobic and hydrophilic, they detergent to serve as digestive juice to emulsify lipid for lipid absorption in the intestinal tract [1]. In 1999, bile acids have been identified as endogenous ligands for nuclear receptor, Farnesoid X receptor (FXR) [5]. In 2006, bile acids have been reported as endogenous ligands for G-protein bile acid receptor (TGR5) [6]. Numerous studies have demonstrated that bile acids promote secretion of glucagon-like peptide 1 (GLP-1) to potentiate insulin secretion in pancreatic β cells, leading to decreased blood glucose level in enteroendocrinal cell lines and animal models [78]. Furthermore, bile acid nuclear receptor FXR has been recently reported to crosstalk with TGR5 to secrete GLP-1 [910]. The discovery of bile acid-mediated endocrine functions of FXR and TGR5 has proposed new perspectives to understand physiological roles of bile receptors to control GLP-1 secretion for the maintenance of glycemic homeostasis.
The biology of bile acids
Abovementioned, bile acids are converted from the cholesterol in the liver by numerous cytochrome P450s. Once bile acids are synthesized, they are immediately conjugated to taurine or glycine for secretion into bile canaliculi [13]. Bile acids are stored in the gall bladder until feeding signal stimulates bile acid secretion for emulsifying lipid and fat absorption in the small intestine. When bile acids reach the small intestine, 95% of secreted bile acids are reabsorbed and transported back to the liver by portal vein. The remaining 5% of secreted bile acids is excreted with feces. Thus, total bile acids pool size loses 5% of bile acids in each enterohepatic circulation. The conversion of cholesterol to bile acids compensates equivalent amount of loss of bile acids excreted with feces. Thus, conversion of bile acid from cholesterol is to maintain constant bile acid pool size [13].
Farnesoid X receptor: a member of nuclear receptor superfamily
Nuclear receptors are well-known transcriptional regulators and regulate diverse biological functions, such as physiological homeostasis, reproduction, development, inflammation and metabolism [11121314]. In general, nuclear receptors function as ligand-activated transcriptional regulator. Upon binding with their endogenous ligands, nuclear receptors recruit various coactivators to induce their target gene expressions. In the absence of their ligands, nuclear receptors interact with corepressor complex to suppress expression of their target genes. Nuclear receptors share common structure. Though the N-terminal domains are highly variable (A/B domain), nuclear receptors share constitutively active transactivation function domain (AF-1). In C-terminal domain, nuclear receptors share ligand-dependent activation function domain (AF2) (Figure 1). The most conserved domain of nuclear receptors is DNA-binding domain (DBD), which has two cysteine-Zn2+ finger motifs to recognize specific DNA elements and allow nuclear receptors binding to the DNA for gene regulation. Besides, DBD domain allows nuclear receptors dimerization to control target gene expression (Figure 1). Ligand-binding domain is the most critical region to control transcriptional activities of nuclear receptors. Upon binding of their ligands, conformational changes of LBD lead to modulation of transcriptional activities of their nuclear receptors (Figure 1). Binding of agonist, the structure changes of LBD recruit coactivators whereas antagonists-bound LBD domain recruits corepressors for their gene expression regulation. Between DBD and LBD, hinge region is located to allow 3D structural changes of nuclear receptors to modulate target gene expressions in response to diverse signals (Figure 1).
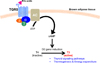
FXR has been identified as a member of nuclear receptor superfamily in 1995 [15]. In the absence of ligands, FXR generally heterodimerizes with retinoid X receptor (RXR) and binds to the FXR response element (FXRE) in the promoter regions with corepressors, including Silencing Mediators of Retinoic acid and Thyroid hormone receptor (SMRT) and histone deacetylases (HDACs) to suppress target gene expression [2151617] (Figure 2). Upon binding with bile acids, FXR interacts with various coactivators including histone acetyltransferase p300, and induces target gene expressions in response to bile acids [18] (Figure 2). FXR is ubiquitously expressed in diverse tissues, such as liver, intestine, kidney, adipose tissues and even immune cells, FXR-mediated bile acid signaling plays pivotal roles to control physiological homeostasis [17192021222324]. Previous studies have reported that FXR-null mice exhibited dysregulated lipid homeostasis [19]. Given that FXR controls a set of genes involved in lipoprotein metabolism including SREBP-1c, PLTP, SCD-1, VLDLR, ApoCII and ApoE, FXR-null mice exhibited elevated cholesterol and triglyceride in the blood [16171925]. Moreover, it has been shown that FXR-null mice exhibited impaired insulin signaling and dysregulated glucose homeostasis [19212224]. Therefore, FXR is a key transcriptional factor to maintain lipid and glucose homeostasis.
Cell membrane bile acid receptor TGR5
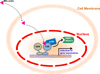
Besides nuclear bile acid receptor FXR, intrinsic bile acid receptor is located in the cell membrane. G protein-coupled bile acid receptor 1 (GPBAR1; TGR5) is a member of the rhodopsin-like superfamily member of GPCR protein and is ubiquitously expressed in diverse tissues, including endocrine organs, muscle, adipose tissue, immune cells, and intestinal tract [82627282930]. Upon binding with lithocholic acids (LCA) and taurolithocholic acid (TLCA), TGR5 is activated to transduce signal transduction into the nucleus and control diverse gene expression [30]. It has been shown that bile acids bind to TGR5 and stimulate cAMP signaling to activate mitogen-activated protein kinase (MAPK) pathway. Elevated intracellular cAMP levels then activate protein kinase A (PKA) to phosphorylate cAMP response element binding protein (CREB) and control their target gene expressions [31]. Previously, TGR5 has been demonstrated to activate thermogenesis and thyroid signaling pathways to enhance energy expenditure in brown adipose tissue [6] (Figure 3). In brown adipose tissue, TGR5 activation induce Dio2 gene expression which converts thyroxine (T4) to tri-iodothyronine (T3), resulting in an increased energy expenditure [6]. In addition to Dio2, numerous genes involved in thermogenesis were largely increased by TGR5 activation: peroxisome proliferator-activated receptor γ coactivator-1α (PCG1α) and 1β (PGC1β), uncoupling protein-1 (UCP1) and -3 (UCP3) and straight-chain acyl-CoA oxidase 1 [6]. Quite interestingly, these gene expressions were not changed in Dio2 knockout mice, suggesting that TGR5-mediated Dio2 functions are critical to induce numerous thermogenic gene expressions [6]. Thus, these findings propose that bile acids control TGR5-cAMP-Dio2-thyroid hormone signaling axis to control energy homeostasis. Consistently, TGR5-deficient mice exhibited severe metabolic syndromes including obesity, severe insulin resistance and impaired glucose and lipid homeostasis [32]. Therefore, these reports clearly have proposed that TGR5 is a potent therapeutic target for the treatment of metabolic syndromes including obesity and type II diabetes.
Bile acids-activated TGR5 to control GLP-1 secretion from the intestinal L cells
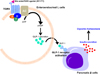
Besides energy expenditure, bile acids have been reported to control incretin secretion, such as glucagon-like peptide 1 (GLP-1). Bile acid-mediated TGR5 activation has been demonstrated to stimulate the secretion of GLP-1, a member of the incretin family [78]. Food intake signals stimulate bile acid secretion from the gall bladder to small intestine, which activates TGR5 in the intestinal entero-endocrine cells to secrete insulinotropic hormones into the bloodstream. In rodent model, bile acids have been shown to activate TGR5 which triggers cAMP signaling pathway to stimulate GLP-1 secretion in intestinal L cells [7]. As consistent, synthetic TGR5 agonist, INT-777 has been shown to potentiate GLP-1 secretion in the intestinal L cells, resulting in GLP-1 receptor activation followed by an increase of insulin secretion in pancreatic β cells [7]. These studies clearly propose that bile acid-mediated TGR5 activation is critical to maintain glucose homeostasis in response to food intake via control of incretin secretion (Figure 4).
Crosstalk between FXR and TGR5 to potentiate GLP-1 secretion from the intestinal L cells
Though bile acid-mediated TGR5 activation on GLP-1 secretion is clear, physiological roles of bile acidmediated FXR activation to control GLP-1 secretion is still controversial. Previous studies have shown that FXR activation repressed transcription of GLP-1 in intestinal L cells [3133]. As gene expression of GLP-1 has been reduced, FXR agonist GW4064 treatment largely reduced GLP-1 secretion in intestinal L cells [33]. In contrast, gut-specific FXR-null mice exhibited elevated GLP-1 secretion, implying that FXR activation is opposite to the TGR5 activation on the control of GLP-1 secretion [33].
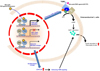
Quite interestingly, another recent study has demonstrated that FXR activation mediated by a gut-specific FXR agonist, Fexaramine stimulates TGR5 expression in the intestinal L cells to potentiate TGR5-mediated cAMP signaling and increases GLP-1 secretion in intestinal L cells [91034]. Consistently, Fexaramine treatment enhanced GLP-1 secretion in genetic obese mice [10]. Moreover, this study has demonstrated that Fexaramine treatment induces gut microbiome remodeling to change bile acid composition and increases the levels of LCA and TLCA which are potent agonists for TGR5. Thus, Fexaramine-mediated microbiome changes lead to change of bile acids composition, resulting in enhanced TGR5 signaling in vivo [10]. Thus, crosstalk between bile acid receptors FXR and TGR5 potentiates GLP-1 secretion in enteroendocrinal L cells to control glucose homeostasis (Figure 5).
Conclusion
It has been largely accepted that bile acid signaling is critical for the beneficial improvements of sleeve gastrectomy surgery in rodent models [35]. Thus, physiological roles of bile acid receptors FXR and TGR5 have been considered as novel therapeutic targets for diverse metabolic syndromes. Consistent with rodent models, there are sufficient evidence suggesting that bile acid signaling has been improved in both human patients and animal models with bariatric surgery [3637]. Patients with beneficial improvements including body weight loss and reduced blood glucose levels exhibited high levels of circulating bile acids and enhanced GLP-1 secretion [363738]. As consistent, patients with severe obesity generally exhibit a decreased postprandial bile acids levels and GLP-1 secretion [38]. Given that crosstalk between FXR and TGR5 is critical to control GLP-1 secretion in response to bile acid signaling pathway, both FXR and TGR5 will provide novel approach to develop therapeutic strategies for treatment of metabolic syndromes such as obesity and type II diabetes.

 XML Download
XML Download