

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Breast cancer is the most common cancer among women worldwide and its incidence has increased in recent years. Breast cancer, with an estimated 1.67 million new cases and 521,900 deaths, accounted for 25% of all cancer cases and 15% of all cancer deaths among females in 2012 [1]. It is thought to be caused by a complex assortment of factors, including personal habits, reproductive factors, social and psychological factors, family history of breast cancer, and genetic factors [2]. Although its pathogenesis is not yet completely clear, researchers generally believe that a variety of environmental factors and genetic factors together lead to the occurrence of breast cancer.
Telomeres, located at the ends of eukaryotic chromosomes, play a key role in the maintenance of chromosome integrity and stability [3]. Telomeres progressively shorten with somatic cell division as a result of incomplete replication at the 3′ end of chromosomes. When telomeres reach a critical threshold, they can lead to chromosomal instability and cell senescence, characterized by apoptosis [4]. Telomere length is maintained by telomerase, a multisubunit ribonucleoprotein polymerase that normally is not expressed in somatic tissue but is elevated in germ cells and neoplastic tissues [5]. When telomerase activity is induced and telomere length altered, cells can escape normal senescence or apoptosis. This leads to cancer cells with unlimited proliferation capacity and the immortalized state [6].
Recently, a series of epidemiological studies have assessed the association between telomere length and risk of various cancers, but the findings remain conflicting [78]. For breast cancer, four studies showed no significant association between telomere length and risk of breast cancer [9101112]; some recent reports have suggested that longer telomere length was associated with increased risk of breast cancer [131415]; however, other investigations indicated that shorter telomere length was associated with increased risk of breast cancer [161718], and one study found that both shorter and longer telomere lengths were associated with increased risk of breast cancer [19]. The reasons for the contradictory results remain unknown. Differences in sample sizes, ethnicity, geography, experimental methods, and background exposures could have contributed to these inconsistent findings.
In addition, previous studies have reported that single nucleotide polymorphisms (SNPs) in the telomerase reverse transcriptase (TERT) gene were associated with susceptibility to breast cancer [15202122]. TERT encodes the reverse transcriptase component of telomerase, which is essential for the maintenance of telomere length, telomerase production, and chromosomal stability [2324]. However, research on the role of TERT genetic variants associated with telomere length is very limited. In this study, we used quantitative real-time polymerase chain reaction (qPCR) to measure relative telomere length (RTL) in genomic DNA extracted from the peripheral blood cells of 183 breast cancer cases and 191 healthy controls, and examined the association between RTL and the risk of breast cancer. Simultaneously, we explored whether these TERT polymorphisms affect telomere length.
Go to : 
METHODS
Study population
This case-control study recruited 183 female breast cancer cases and 191 healthy controls. The cases were diagnosed with breast cancer between January 2013 and November 2015, at the Affiliated Hospital of Xi'an Jiao Tong University, Xi'an, China. There were no age, histological, or tumor stage restrictions in this study. However, patients that had other types of cancers, or who underwent radiotherapy or chemotherapy were excluded from this study. The control subjects without any history of cancer or other medical diseases were randomly selected from healthy volunteers who visited the health examination center at the hospital during the same period. All control subjects were unrelated ethnic Han Chinese women and their ancestors had lived in the region for at least three generations.
The clinical and demographic characteristics of all enrolled subjects, including region, ethnicity, age, height, weight, menopause status, clinical stage, tumor size, tumor estrogen receptor status, tumor progesterone receptor status, lymph node metastasis, primiparous age, age of menarche, and breastfeeding duration were collected using a detailed questionnaire conducted by trained interviewers, as shown in Table 1.
All procedures performed in studies involving human participants were in accordance with the ethical standards of the Ethics Committee on Use of Human Subjects of the Affiliated Hospital of Xi'an Jiao Tong University (2013-36) and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. Written informed consent was obtained from all participants before participation in the study.
DNA extraction
Peripheral venous blood (5 mL) was collected from each participant using vacutainer tubes containing ethylenediamine tetraacetic acid, and stored in a freezer at −20℃ for DNA extraction. Genomic DNA was extracted from whole blood using the GoldMag-Mini Whole Blood Genomic DNA Purification Kit (GoldMag Co., Ltd., Xi'an, China) according to the manufacturer's instructions. The DNA samples were preserved at −4℃ for future use. DNA was quantified using a spectrophotometer (NanoDrop 2000; Thermo Fisher Scientific, Waltham, USA) to ensure accurate and uniform concentration.
Single nucleotide polymorphism selection and genotyping
We selected three polymorphisms in TERT (rs2853677, rs2242652, and rs10069690) previously reported to be associated with breast cancer susceptibility with a minor allele frequency >0.05 for further genotyping. The amplification and extension primers for SNPs were designed using the Sequenom MassARRAY® Assay Design 3.0 Software (Sequenom, San Diego, USA). Genotyping was performed using the Sequenom MassARRAY® platform according to the standard protocol recommended by the manufacturer. Sequenom Typer 4.0 software was used to perform data management and analysis.
Relative telomere length measurement
Telomere length was measured using the real-time qPCR method as described by Cawthon [25]. The relative ratio of the telomere (T) repeat copy number to a single gene (S) copy number (T/S ratio) was established for experimental samples using standard curves. The ratio for each sample was normalized to a reference DNA to standardize between different runs. 36B4 on chromosome 12, encoding acidic ribosomal phosphoprotein P0, was used as the single copy gene. All samples for both the telomere and 36B4 gene amplifications were performed in duplicate in separate 96-well plates. The cycle threshold (Ct) is the number of cycles required for the fluorescent signal to cross the threshold. Ct values generated were used to calculate the T/S ratio for each sample using the equation: T/S=2−ΔCt, (ΔCt=Cttelomere sample−Ct36B4). Telomere length was expressed as a relative T/S ratio normalized to the average T/S ratio of the reference sample [2−(ΔCtx−ΔCtr)=2−ΔΔCt].
Statistical analysis
We used Pearson chi-square test and Welch t-test to examine differences of categorical variables and continuous variables between cases and controls, respectively. The Mann-Whitney U-test was used for RTL comparison. The association between RTL and breast cancer risk was estimated by odds ratio (OR) and 95% confidence interval (CI) using the logistic regression model with adjustments. A restricted cubic spline curve was plotted in the logistic regression model to evaluate the shape of the association between RTL and risk of breast cancer. All statistical tests were two-sided, and p<0.05 was considered statistically significant. Statistical analyses were performed using Microsoft Excel (Microsoft Corp., Redmond, USA) and SPSS version 19.0 software (IBM Corp., Armonk, USA).
Go to :
RESULTS
Characteristics of the study population
Table 1 summarizes the demographic characteristics and clinical information of the study participants. The mean age was 46.40±9.38 years in the case group and 45.35±6.90 years in the control group. The mean body mass index (BMI) was 23.08±3.00 kg/m2 in the case group and 22.53±2.55 kg/m2 in the control group. Welch t-test showed no statistical difference between case and control groups for mean age and BMI distribution (p=0.218 and p=0.056, respectively). Pearson chi-square test showed no statistical difference between case and control groups for the menopause status distribution (p=0.716).
Relative telomere length distribution between case and control groups
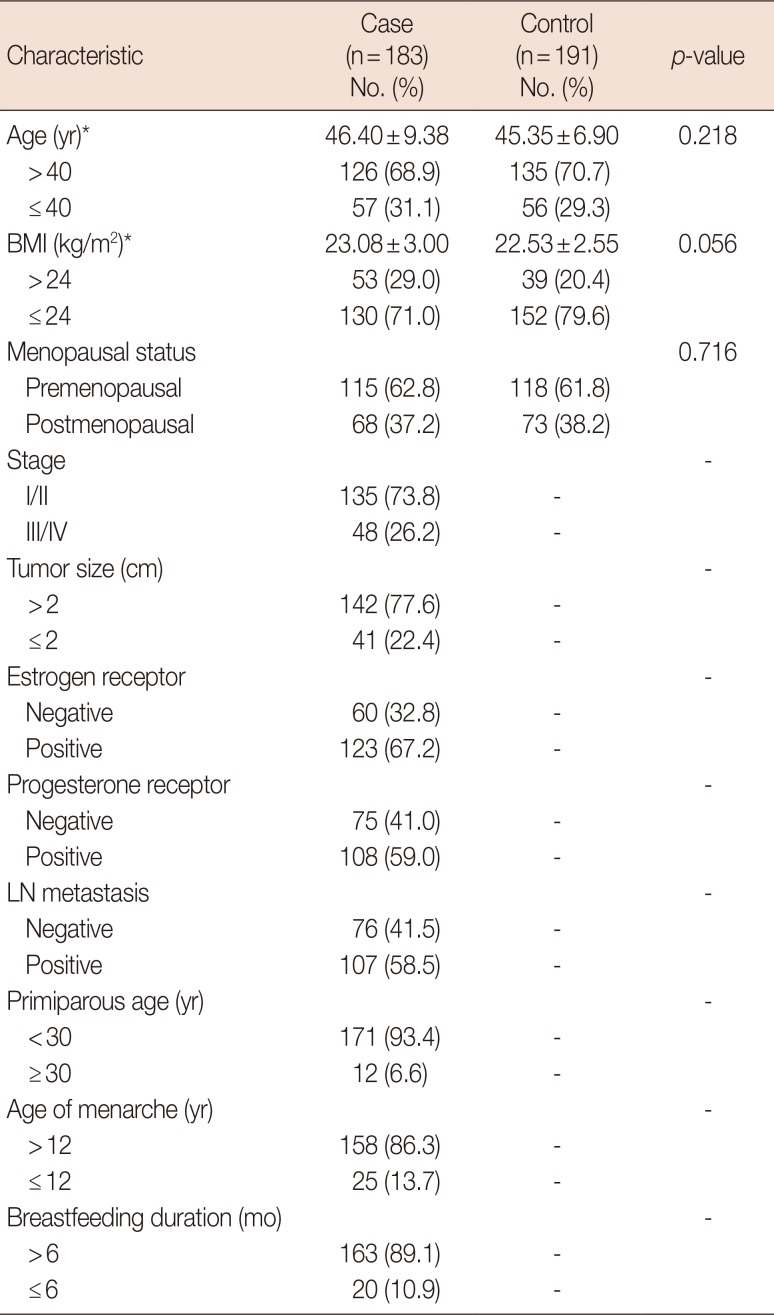
The total mean RTL was 0.824 in the control group and 0.640 in the case group. Breast cancer patients had significantly shorter RTLs than control subjects, as shown in Figure 1 (p=0.001). The age of 40 years, BMI of 24 kg/m2, and menopause status were used for stratification and Mann-Whitney U-test showed that the mean RTL distribution was statistically different between case and control groups at age >40 years, BMI ≤24 kg/m2, premenopausal, and postmenopausal (p=0.003, p<0.001, p=0.045, and p=0.003, respectively). However, no significant differences in the distribution of mean RTL were found between case and control groups at age ≤40 years and BMI >24 kg/m2, age ≤40 years and age >40 years, BMI ≤24 kg/m2 and BMI >24 kg/m2, premenopausal, and postmenopausal in breast cancer patients and healthy control groups (p>0.05), as shown in Figure 1.
 | Figure 1Mean relative telomere length (RTL) in breast cancer cases and healthy controls. (A) Reduced mean RTL of breast cancer cases (n=183) compared to controls (n=191). The distribution of mean RTL between case and control groups at the subgroup level for age (B), body mass index (BMI) (C), and menopause status (D). Horizontal lines represent the mean RTL; p-values were calculated using the Mann-Whitney U-test.
|
Association of relative telomere length with breast cancer risk
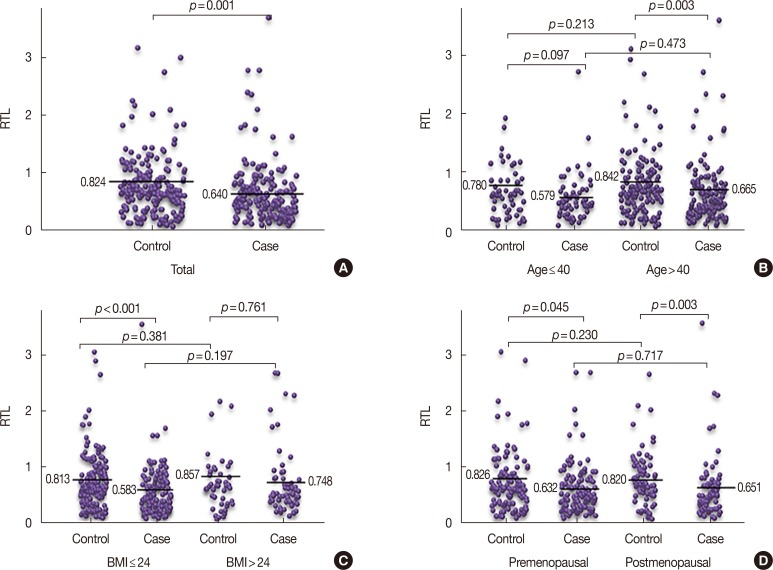
Next, we investigated the association between RTL and breast cancer risk using logistic regression analysis (Table 2). The subjects were categorized according to the mean RTL, and we observed a statistically significant increased risk of breast cancer associated with shorter telomere length (≤0.61) compared with longer RTL (>0.61) after adjusting for age, BMI, and menopause status (OR, 2.13; 95% CI, 1.40–3.23; p<0.001). When the RTL was categorized into tertiles, we found that the lowest RTL (<0.44) was also significantly associated with increased breast cancer risk compared with the highest RTL (>0.77) after adjusting for age, BMI, and menopause status (OR, 2.33; 95% CI, 1.40–3.90; p=0.001). The results of the restricted cubic spline curve analyses indicate that shortened telomere length was associated with breast cancer risk (Figure 2).
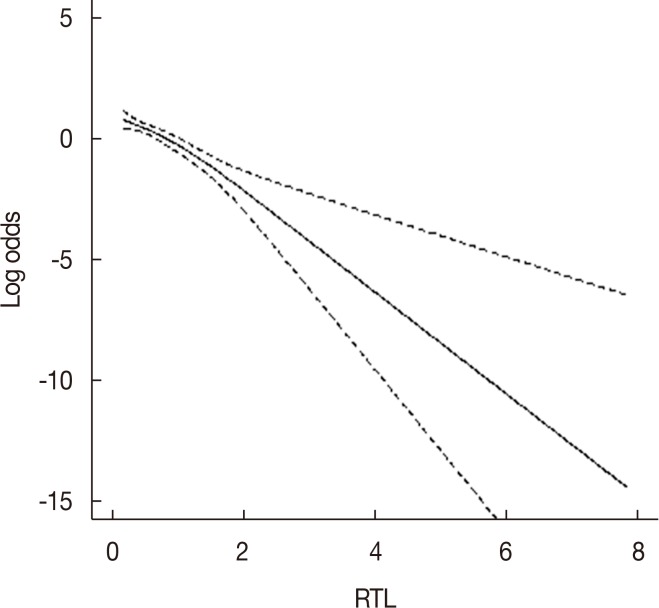
We also performed subgroup analyses by age (≤40 years and >40 years), BMI (≤24 kg/m2 and >24 kg/m2), and menopause status (premenopausal and postmenopausal) as shown in Table 2. The results indicate that the lowest RTL was significantly associated with an increased risk of breast cancer in comparison with the highest RTL in age >40 years after adjusting for BMI and menopause status (OR, 2.41; 95% CI, 1.31–4.43; p=0.005). In BMI ≤24 kg/m2, the lowest RTL was significantly associated with increased risk of breast cancer after adjusting for age and menopause status (OR, 2.81; 95% CI, 1.55–5.10; p=0.001). Increased breast cancer risk for the lowest RTL was also noted in postmenopausal women compared with the highest RTL after adjusting for age and BMI (OR, 3.94; 95% CI, 1.63–9.51; p=0.002).
TERT polymorphism genotypes and relative telomere length
To test the hypothesis that telomere length may be compromised by the TERT SNPs rs2242652, rs10069690 and rs-2853677, which were previously reported to be associated with breast cancer risk, we analyzed their association with RTL in 183 breast cancer patients according to TERT polymorphism genotype distribution. We found that carriers of the genotype AA of rs2853677 had longer telomeres than individuals with the AG genotype (p=0.011), as shown in Figure 3. However, there was no association between the rs2242652 and rs-10069690 genotypes and RTL in the case group (p>0.05).
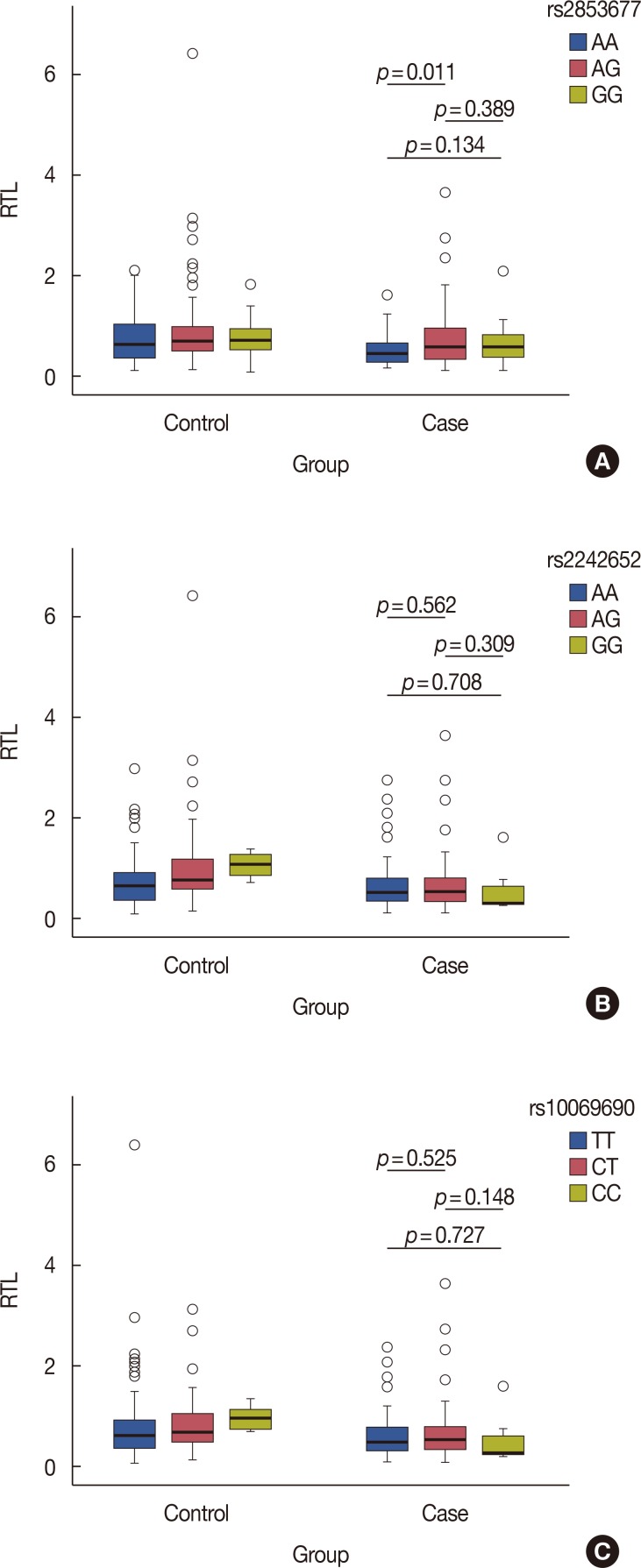 | Figure 3Relative telomere length (RTL) in 191 controls and 183 breast cancer patients according to telomerase reverse transcriptase polymorphisms genotypes distribution. (A) rs2242652; (B) rs10069690; (C) rs2853677. Boxes represent the interquartile range of distributions (25th–75th percentile); horizontal lines within the boxes represent the medians; vertical lines represent the 5th and 95th percentiles; p-values were calculated using chi-square test (two-sided). p<0.05 indicates statistical significance.
|
Go to :
DISCUSSION
In the present case-control study with 183 breast cancer cases and 191 unrelated healthy female controls, we observed that shorter RTL was associated with an increased risk of breast cancer. The subgroup analyses show that risk of breast cancer was significantly increased in the lowest RTL in comparison with the highest RTL in age >40 years, BMI ≤24 kg/m2, and postmenopausal women. Breast cancer patients with the AA genotype of rs2853677 had longer telomeres than individuals with the AG genotype. However, no association between the genotypes of rs2242652 and rs10069690 and RTL was found in breast cancer patients.
Several studies have examined the association between telomere length and breast cancer risk, with contradictory results. Barwell et al. [9] confirmed that there was no significant difference between breast cancer patients and controls in the mean overall telomere length in blood leukocytes. A prospective study showed that RTL was not associated with a significant elevation breast cancer risk in postmenopausal women [10]. Zheng et al. [11] reported that overall telomere length in blood leukocytes was not significantly associated with the risk of breast cancer. Results of a prospective case-cohort analysis from the Sister Study showed that there was also no significant association between RTL in peripheral blood cells and breast cancer risk [12].
Several studies have demonstrated an association of longer telomere length with breast cancer risk. In a case-control study of 265 newly diagnosed breast cancer patients and 446 female controls, Svenson et al. [13] indicated that the case group displayed significantly longer telomere length compared with the control group, and longer overall telomere length in peripheral blood leukocytes was strongly associated with increased breast cancer risk. Gramatges et al. [14] found that breast cancer cases had significantly longer RTLs compared with the control group. Pellatt et al. [15] also observed that longer telomere length was associated with increased breast cancer risk (OR, 1.87), with the highest risk among women with high Indigenous American ancestry (OR, 3.11). The assessment of risk by RTL quartile showed an increased risk for breast cancer with each longer quartile, with the most significant risk observed in the longest quartile in a Stanford population.
Shen et al. [16] observed that overall telomere length was not significantly associated with breast cancer risk in all subjects. However, they noted that shorter telomere length may be associated with an increased risk of breast cancer in premenopausal women. The results also showed that presence of shorter telomeres was associated with a significantly increased breast cancer risk in premenopausal women [17]. Pooley et al. [18] found a strong association between shorter telomere length and breast cancer risk in a retrospective study. Our results suggest that shorter RTL in blood cells was significantly associated with breast cancer risk in all subjects. The subgroup analyses showed that shorter telomere length was also associated with an increased risk of breast cancer in age >40 years, BMI ≤24 kg/m2, and postmenopausal women. A large case-control study revealed a reverse J-shaped association between telomere length and breast cancer risk, suggesting that shorter or longer telomeres were associated with an elevated risk of breast cancer [19]. Results from these studies have been inconsistent, perhaps because of the differences in sample size, race, geography of study population, and experimental methods.
Although the mechanisms underlying breast cancer pathogenesis are not fully understood, it has been proposed that telomeres progressively shorten with ongoing cell division until they reach a critical length that induces replicative senescence, leading to a concurrent increase in genomic and chromosome instability, thereby increasing susceptibility to cancer [26]. Other researchers have reported that telomerase expression prevents further telomere shortening correlated with the capacity for unlimited cell proliferation in malignant tumors, thus increasing the risk of cancer [27]. Additional research is needed to explore the relationship between telomere length and breast cancer pathogenesis and the underlying mechanisms.
The carriers of the AA genotype of rs2853677 had longer RTLs than individuals with the AG genotype in the case group. This finding supports the observation that carrying the rs2853677 T-allele reduced breast cancer-specific mortality (hazard ratio, 0.57; 95% CI, 0.39–0.84) [28]. There was no association between the genotypes of rs2242652 and rs10069690 and RTL in breast cancer patients. These findings suggest that rs10069690 and rs2242652 may increase the risk of breast cancer through mechanisms other than telomere length. Genotypes of rs10069690 did not show any association with mean RTL in cases of paranoid schizophrenia or controls [29]. However, longer RTL was observed among carriers of rs10069690 and rs2242652 genotypes in a Utah population [30]. Further studies are warranted to validate this association and investigate our findings in a larger sample size.
Some limitations of this case-control study should be considered when interpreting the results. First, the sample size of this study comprising a total of 183 breast cancer cases and 191 unrelated healthy controls was relatively small, so it is necessary to expand the sample size to confirm the results. Second, our results are limited to Chinese Han women, and further studies are required in other populations. Third, other risk factors such as smoking or alcohol consumption could not be evaluated because of lack of data. Fourth, the association of TERT polymorphisms with breast cancer risk was not verified in this study. In addition, biological functional analyses of TERT polymorphisms were not performed.
In conclusion, this study suggests that shorter RTL was associated with an increased risk of breast cancer in all subjects, age >40 years, BMI ≤24 kg/m2, and postmenopausal women. Breast cancer patients with the genotype AA of rs2853677 had longer RTLs than patients with the AG genotype. No association was found between the geno types of rs2242652 and rs10069690 and RTL. The results suggest that rs2853677 reduces the all-cause mortality of breast cancer through longer telomere length, while rs10069690 and rs2242652 may increase the risk of breast cancer through mechanisms other than telomere length. The results support an important role of telomere length as a biomarker of breast cancer risk. Functional studies are warranted to validate this association and further investigate our findings in a larger sample size.
Go to :
 XML Download
XML Download