

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Cancer remains the most important health issue of the day. It is estimated that more than 26 million people will be diagnosed with cancer, resulting in 13 millions death in 2030 [1].
Cancer frequency is increasing by 1% to 2% per year in almost every country in the world. Breast cancer (BC) accounts for approximately 30% of female cancers in the United States and approximately 25% of all cancers worldwide. The risk of devoloping BC during her lifetime is an estimated 10% to 12.8% [12].
Biological factors primarily associated with BC are the estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor 2 (HER2), and proliferation. Triple-negative breast cancer (TNBC) is characterized by ER, PR, and HER2 expression deficiency. Studies aimed at finding a tumor marker specific for BC have been continuing for many years. However, no biochemical or immunochemical substance that can be quantified has been determined yet [3].
The main methods used in BC therapy are surgery, radiotherapy, chemotherapy, hormone therapy, and targeted therapy [4]. The most important goal of developing any effective cancer treatment method must be the exclusive destruction of tumor cells while preventing cancer recurrence. Immunotherapy is a new generation treatment method developed to overcome side effects of conventional treatments while increasing effectiveness by providing tumor-specific treatment via targeting of tumor cells with antigen differences [5].
Owing to the weak antigenicity of antigenic structures in cancer cells, cancerous cells cannot be detected via immunosurveillance or eliminated by immunocytes, which allows the tumor to develop unhindered [6].
Immunotherapy is an approach that stimulates immune system cells and renders them more effective. In addition to supporting the immune sysrem, the wider goal of immunotherapy is to allow immune system cells to recognize tumor antigens more effectively and specifically attack and destroy cancer cells [7].
Go to : 
GENERAL APPROACHES ON VACCINE DEVELOPMENT AGAINST BREAST CANCER
Peptide-based vaccines
In order to develop an effective therapeutic vaccine against BC, self-antigens of breast tumor cells that serve as tumor targets are used. The HER2 protein, specific for both trastuzumab and pertuzumab monoclonal antibodies, was the first antigen to be identified as a target for BC immunotherapy. HER2, carbohydrate antigens (such as carcinoembryonic antigen), and Mucin-1 (MUC-1) are the most commonly studied BC antigens for the purpose of vaccine formulation. However, antibody response to HER2 and MUC-1 antigens in BC patients is very low. Peptide vaccines have been used mostly with the granulocyte macrophage colony stimulating factor (GM-CSF) cytokine adjuvant to enhance efficacy [8].
Of the different combinations that may target MUC1 tumor associated antigen (TAA), the BLP25 MUC1 lipopeptide-containing mixture has been used against BC, in addition to chemotherapy and showed an increase in CD8+ T responses. Currently, the possibility of using the MUC1 antigen in vaccine formulation is being tested in many clinical trials.
Bacille Calmette-Guérin (BCG) conjugated with MUC1 antigen and human interleukin-2 (IL-2) vaccine caused an increase in the cytotoxic T lymphocyte (CTL) response in mouse models [8]. This vaccination approach has been shown to be more effective than the formulation containing the polyinosinicpolycytidylic (poly-IC) stabilized with poly-lysine and carboxymethylcellulose (poly-ICLC) adjuvant [9].
Adjuvants and helper peptides have been used to improve the clinical efficacy of peptide vaccines. An example of this approach is study conducted by Dillon et al. [10], using the major histocompatibility complex class I-restricted BC-associated and Toll-like receptor 3 agonist poly-ICLC adjuvant.
Mittendorf et al. [11] demonstrated the efficacy of the HER2/neu derived peptide GP2 with nine amino acids (654–662: IISAVVGIL) peptide vaccine in preventing tumor recurrence, when GP2 is used together with the GM-CSF adjuvant.
Torres-García et al. [12] evaluated parasite-derived immunomodulatory 18-amino acid (aa) GK-1 peptide adjuvant in vivo and reported an enhanced inhibitory effect on invasive BC and its metastasis to lungs.
Currently, NeuVax (Galena Biopharma, San Ramon, USA), and Pan-T-cell epitope+a neolactoseries antigen Lewis Y+the ganglioside GD2 mimicking carbohydrate peptide containing vaccine P10s-PADRE (AmbioPharm Inc., North Augusta, USA), are being evaluated in clinical trials for use against BC. Furhter examples of clinical trials on peptide vaccines are shown in Table 1 [1013141516171819].
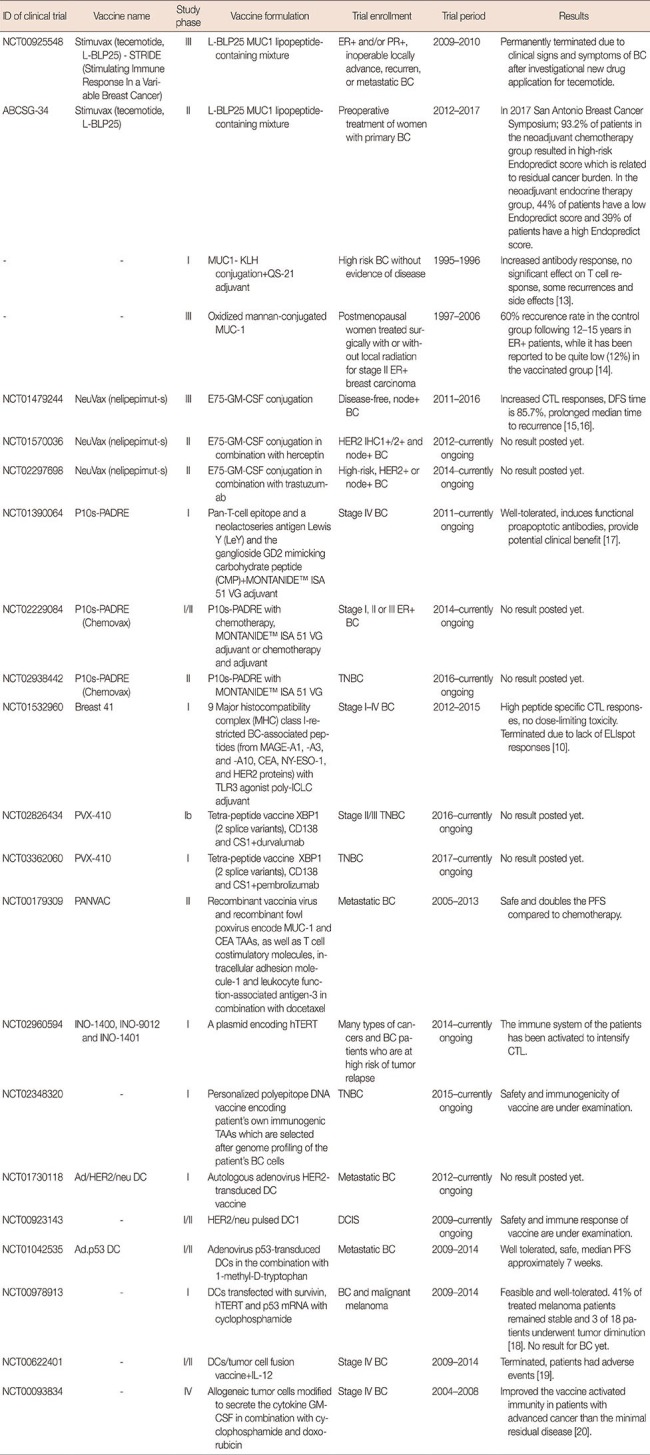
Table 1
Clinical trials on breast cancer vaccines
MUC1=Mucin-1; ER=estrogen receptor; PR=progesterone receptor; BC=breast cancer; KLH=keyhole limpet hemocyanin; GM-CSF=granulocyte macrophage colony stimulating factor; CTL=cytotoxic T lymphocyte; DFS=disease-free survival; HER2=human epidermal growth factor receptor 2; IHC=immunohistochemistry; TNBC=triple-negative breast cancer; CEA=carcinoembryonic antigen; TLR3=Toll-like receptor 3; poly-ICLC=carboxymethylcellulose, polyinosinic-polycytidylic acid, and poly-L-lysine double-stranded RNA; TAA=tumor associated antigen; DC=dendritic cell; hTERT=human telomerase reverse transcriptase; DCIS=ductal carcinoma in situ; PFS=progression-free survival; IL-12=interleukin 12.
![]()
Whole-cell vaccines
Vaccine platforms based on direct application of whole tumor cells or cell extracts to the patient, as an alternative to antigen-specific vaccination with peptide or protein subgroups, are direct polyvalent immunization strategies [20]. Whole-cell tumor vaccines present multiple and unknown cancer antigens in contrast to epitope-specific vaccines. However, according to current research these vaccines are poorly immunogenic [21].
Whole-cell tumor vaccines are prepared by irradiating allogeneic or autologous cancer cells or cell lysates, which inhibits their ability to replicate inside the patient following vaccination [22]. Irradiating cancer cells enhances tumor recognition by CTLs and antigen-presenting cells [23].
Certain modifications have been made in order to increase vaccine immunogenicity. The GVAX (Aduro Biotech, Berkeley, USA) vaccine includes irradiated allogeneic tumor cell lines which are modified to secrete the cytokine GM-CSF [24]. The potency of the GM-CSF secreting whole cell BC vaccine can be enhanced by using chemotherapeutic agents.
Huang et al. [25] added IL-2 cytokine and blocked immunosuppressive anionic phospholipid phosphatidylserine in the whole cell cancer vaccine to increase its efficiency. An increase in the tumor-specific cytotoxicity response and interferon (IFN)-γ cytokine release has been observed in vaccinated mice. Convit et al. [26] prepared a vaccine formulation including whole-tumor cells, formalin, and BCG adjuvant to improve vaccine immunogenicity and proved the tolerability, safety, and nontoxicity of the formulation based on the results of animal research. The results of this study indicated an overall survival of 60% in humans. The vaccine formulation resulted in a lower tumor growth rate and boosted immune cell activation in the mouse model. Promising results in the term of a decrease in the infiltration of suppressor cells and improved immune memory have been observed [27].
Immunotherapy with vascular endothelial growth factor receptor 2 (VEGFR2) model antigen showed great potential as a tumor vaccine. Researchers irradiated the adenovirus encoding VEGFR2 (AdVEGFR2)-infected whole cancer cell vaccine-based immunotherapy to improve its efficacy. The results of this study indicate that angiogenesis inhibition, CTL activation and increased expression of the high mobility group box 1 (HMGB1) and 70 kilodalton heat shock proteins in AdVEGFR2-infected cells may be used as an effective strategy in cancer immunotherapy [28].
MDA-MB-231 cell line has been used as a whole tumor cell vaccine in a stage IV BC clinical trial. This cell line is an HLA-A21, HER2/neu1 allogeneic BC cell line, which was genetically modified to express the costimulatory molecule CD80 (B7-1). This formulation was used with GM-CSF and BCG adjuvant, resulting in a costimulatory signal and improved antigen presentation capability in patient T cells. It has been proved that the vaccine formulation is safe and feasible [29]. Furhter examples of clinical trials on whole cell vaccines are shown in Table 1.
Although using tumor cell antigens has improved the efficacy of these applications, the inability of recognition of immunocompetent agents, which are suppressed by tumor cells, to recognize the immunogens may be one of the greatest reasons underlying the inability of these vaccine formulations to achieve the desired activity.
Gene-based vaccines
In recent years, innovative gene-based vaccines have also been developed for use in treating BC. Recombinant viral vector vaccines based on New York Vaccinia, modified virus of Ankara (MVA) and canarypox virus have been designed to express TAAs such as HER2, p53, and MUC1, and have been generally tested in clinical trials involving metastatic BC patients [30].
Kwilas et al. [31] designed a poxviral-based cancer vaccine using MVA including the Twist transgene and a triad of costimulatory molecules (B7-1, intercellular adhesion molecule 1, lymphocyte function-associated antigen 3) and evaluated its efficiency in metastatic BC and prostate cancer models. They targeted the Twist transcription factor which plays an important role in metastasis, poor prognosis, and drug resistance, and observed both CD4+ and CD8+ Twist-specific T-cell responses in vivo.
Although these viral vaccines produce highly potent CTL responses to encoded TAAs, they pose a risk in clinical practice because of the potential for inducing strong immunity to the viral construct and, in some cases, neutralizing antibodies. In addition, a major issue preventing intensive use of the viral vector-based vaccines is the high level of immune response to viral antigens as compared to coded antigens. A similar issue exists in adenovirus-based vaccines. Elimination of certain high immunogenic motifs such as the E1a and E2b genes may attenuate antibody neutralization after vaccination [32].
However, development of DNA vaccines has afforded a new perspective to cancer immunotherapy. The approval of Oncept™ (Merial, Duluth, USA), a plasmid coding for human tyrosinase, by Food and Drug Administration for canine melanoma proved the importance of DNA vaccines in 2010 [33].
Nazarkina et al. [34] designed a polyepitope DNA vaccine encoding immunogenic peptides of HER2 and mammaglobin-1 tumor antigens and optimized its delivery to dendritic cell (DC). It was demonstrated that secretion of IL-6 from DNA vaccine transfected DC, indicating DC maturation, and has great capacity to induce an immune response. Furhter examples of clinical trials on whole cell vaccines are shown in Table 1.
Although promising results have been obtained using recombinant vaccines in BC immunotherapy, a major risk associated with the use of these vaccine formulations is the possibility of oncogene activation and stimulation of cancer, as well as the possibility that genetic material in the vaccine may fuse with the genome of healthy cells in the patient.
Dendritic cell-based vaccines
One of the most important approaches developed for treating BC is the DC-based immunotherapeutic vaccine. DC-mediated vaccine development approaches against BC are based on preparing synthetic peptides in accordance with the structure of antigens on BC cells, and obtaining immunogenic antigens by killing autologous or allogeneic BC cells using various means such as gamma irradiation, chemical sensitization, treatment with chemotherapeutic drugs or freez-thaw-based techniques and loading the isolated antigens in vitro to DCs. In addition, tumor cells may be fused with DCs using polyethylene glycol (PEG) or electrofusion technology, thereby providing tumor antigen presence on DCs [35].
Her2/neu and MUC1 are peptide antigens that are widely used in DC vaccines are. Lapuleucel-T (APC8024), overexpressing HER2 peptide, which has been developed for use in cancer patients and tested in phase I clinical trials, is obtained via the recombinant fusion protein BA7072 in DCs of peripheral blood mononuclear cells (PBMC) origin in vitro [36]. This fusion protein is a product of the conjugation of intracellular and extracellular domains of HER2 with the GM-CSF adjuvant. This study proved the safety and anti-HER2 effect of Lapuleucel-T. It also indicated that this formulation may cause less side effects compared to Sipuleucel-T vaccine [36].
However, since such vaccination approaches can only be used in BC patients overexpressing HER2 or MUC-1 peptides, their efficacy for use in other types of BC that do not express these peptides or do not have a specific antigen, such as TNBC, is considered to be insufficient.
Additionally, certain studies using genetically modified DCs have been reported. Sakai et al. [37] used recombinant adenovirus-transduced bone marrow-derived DCs (BMDCs) encoding neuroproteins in the vaccination of the BALB-neuT BC mouse models. The vaccine has shown positive results such as specific anti-neu antibody production, improved interferon gamma release, and delayed carcinoma development in mice. However, this study is valid only for HER2/neu-expressing BCs and is considered to be inadequate for other BC cell.
Other studies involve the use of all tumor antigens in order to remove limitations caused by utilizing single or multiple antigens in DC-based immunotherapeutic approaches. These studies are based on the fusion of tumor cells and DCs (PEG and electrofusion), or loading of tumor cells into DCs using various techniques for obtaining antigens (gamma irradiation, treatment with chemotherapeutic drugs, and so forth).
In one such study, Gong et al. [38] demonstrated that lysis of cancer cells may be achieved by evaluating in vitro CTL response of hybrid cells formed via performing PBMC-based DC-tumor cell fusion PEG. In 2007, Koido et al. [39] performed PEG-mediated fusion with DCs of MCF-7 and BT-20 tumor cells and estimated the efficacy of this vaccine both in vitro and in a mouse model in vivo. Although in vitro studies have shown high CTL responses, the main disadvantage of this study is that the vaccine prepared using human BC cell lines, was tested only in mouse cancer models. This prevents achievement of reliable results. Bird et al. tested PEG fusion-based DC vaccines in dog models in two different studies in 2008 and 2010. The former involved the fusion of DCs with allogeneic cancer cells [40] whereas the latter involved fusion with autologous cancer cells [41]. Effective CTL responses were not observed in these studies. It is proposed that DC vaccines based on PEG fusion are different from DC function and antigen presentation, and that vaccine based on such formula may suppress immune responses by activating the Treg cells.
Interest in electrofusion technology in the production of DC-based vaccines has increased recently owing to its usefulnuss in avoiding toxic effects of PEG fusion as well as increasing the efficiency of fusion. Zhang et al. [42] performed the fusion of DCs with MCF-7 BC cell line using an electrofusion device. Analysis of the immune system-stimulating effect of in vitro generated hybrid cells, determined that, in addition to high T lymphocyte proliferation and cytokine release responses, MCF-7-specific CTL responses could also be obtained. However, research has indicated that DC-based vaccines generated by allogeneic tumor cells in this manner, may only be useful for vaccination protocols on an experimental basis.
In another study, autologous BC cells were electrofused to autologous DCs, where it was reported that formulations in which other products of the fusion mixture were used, demonstrated a more effective immunostimulatory effect than the ones using purified hybrid cells [43]. Zhang et al. [44] performed electrofusion of PBMC-derived DCs with MDA-MB-231 cell lines, evaluated in vitro T cell proliferation, IL-12 and IFN-γ release, and tumor-specific CTL responses, and reported more effective responses compared to control groups. The results of this study showed that fusion with of DCs with whole BC cells produced an effective cancer specific antitumor T cell response in vitro, which made such a procedure a promising candidate for use in adoptive immunotherapy. Studies indicate that immunotherapeutic dendritic/cancer cell fusion hybrids may inhibit the induction of CTL responses due to soluble factors derived from tumor cells and that the formation of immunosuppressive regulatory T cells is supported [45].
Isolation of antigens from cancer cells, used as an antigen source, to be loaded to DCs can be achieved in a similar manner as Neidhardt-Berard et al. [46], who performed MCF-7 cell line culturing in serum-free medium and gamma irradiation and HCC1806 cell line anti-Fas monoclonal antibody, CH11 and cycloheximide desensitization in the form of killing. The researchers tested in vitro CTL responses and IFN-γ cytokine release by loading the antigens to DCs and enabling those with T lymphocytes. While this approach demonstrated the efficacy of stimulating immunosystem cells, it also emphasized that different methods of antigen isolation may be necessary since the killing methods used, did not demonstrate similar levels of efficacy in all BC cells. In a study conducted by Saito et al. [47], T47D and Hs578T cell lines were been killed using betulinic acid. The researchers induced CTL formation by culturing the tumor lysates with DCs and T lymphocytes. Cyclin B1-MUC1- and survivin-specific CTL responses were determined in vitro. However, betulinic acid killing is not a commonly reported technique in the literature.
Delirezh et al. [48] determined the immunostimulatory efficacy of DCs loaded with gamma γ irradiation killed human autologous BC cells in vitro. Antigen-loaded DCs were cultured with autologous T lymphocytes from patients, resulting in high T cell proliferation and IFN-γ cytokine production response. Individual-specific CTL response emphasizes the importance of autologous cell use in the vaccine protocol. However, the gamma irradiation based killing method is not effective in killing every cancer cell.
The freeze-thaw method, which causes instantaneous temperature changes in the cell leading to isolate of whole antigens, is a method that has been proved reliable for many. This method is often used in anticancer studies because of the low number of antigens identified on cancer cells and the ability to release certain idiotypic epitopes or antigens, which are products of mutational events occurring throughout the tumorigenic process in the cancerous cells [49].
The heat shock soluble proteins obtained by the freeze-thaw method increased antigen presentation and expression of helper stimulating molecules in DCs [50]. As an example of this method, the in vitro and in vivo immunotherapeutic efficacy of a vaccine formulation prepared by loading freeze-thawed antigens of BC cells onto DCs was evaluated by Delirezh et al. [51]. The study concluded that both CD4+ and CD8+ T lymphocyte-mediated immunity was acquired in vitro.
Another immunotherapeutic approach in the treatment of BC is a vaccination involving loading of DCs with antigens of cancer stem cells, which are the main factors that cause inadequacies in traditional treatments. Although only a few studies using this approach have been reported, it may be beneficial both in preventing recurrence as well as metastasis of cancer stem cells, and in providing immunosuppression by permanent memory cells in case the resting cancer stem cells become active.
Nguyen et al. [52] assessed the specificity of DCs developed to overcome cancer stem cells both in vitro and in vivo. 4T1 tumor cells from BALB/c females were developed using the verapamil-resistance selection method and their antigens collected via freeze-thaw method were loaded onto mouse BMDCs. This improved vaccine formulation stimulated breast cancer stem cell (BCSC)-specific CTL responses in in vitro studies. Improvements were also observed in tumor size, as indicated by a decrease in the tumor size of the treated group in vivo in tumor-induced Balb/c mouse models of BCSC tumors. These results demonstrated that DCs may be used in cancer stem cells as well as in the treatment of drug-resistant cancer cells.
Following demonstration of the importance of DC vaccines in BC immunotherapy in both in vitro and in vivo studies, clinical trials in this field as well as studies on DC vaccines have increased dramatically. Detailed explanations of these studies are presented in Table 1.
Nanotechnology on breast cancer vaccines
Recent developments in the field of nanotechnology show a great promise in providing solutions to the insufficiencies currently affecting the development of effective vaccines against BC. Nanoparticles (NPs) confer many advantages to vaccine technology such as increased circulation time, immunogenicity, biocompatibility and solubility of hydrophobic antigens, controlled as well as targeted release and protection of peptide antigens from enzymatic degradation. Nanosized particles in the 10–50 nm are able to easily reach DCs in the lymphatic system. Also, larger NPs are mostly captured by macrophages. Thus nano-designed vaccine formulations can boost immune system components much more effectively [53].
Polymer-peptide conjugates are one of the most common nano delivery systems used in vaccine development. Razazan et al. [54] conjugated GP2, a HER2/neu-derived peptide, to distearoylphosphoethanolamine-N-(maleimide[polyethylene glycol]-2000) micelles. Next, the peptide, including micelle, was inserted into liposomes composed of (dimyristoylphosphatidylcholine [DMPC], dimyristoylphosphoglycerol [DMPG]) phospholipids, and fusogenic lipid dioleoyl phosphatidylethanolamine (DOPE) containing monophosphoryl lipid A (MPL) adjuvant (DMPC-DMPG-DOPE-MPL-GP2). The researchers have evaluated the immune response of BALB/C mice immunized with the vaccine formulation. Liposome-DOPE-MPL-GP2 vaccine formulation produced the high level of IFN-γ release and increased CTL responses. The prophylactic effect of liposome-DOPE-MPL-GP2 was determined as a mean tumor growth delay of 92.22%.
Similarly, Arab et al. [55] prepared E75 antigene attached liposomes consisting of distearoylphosphocholine/distearoyl-phosphoglycerol/cholesterol (Chol)/DOPE nanovaccine to improve immunogenicity of the peptide and evaluated immune responses in vaccinated mice. The results of this study indicated that the vaccine formulation had dramatically increased the CTL response. The significantly prolonged survival time was indicated as a mean tumor growth delay of 72.4%.
Another nano carrying system for peptide vaccines was designed as the AE36 HER2/neu-derived peptide with or without the CpG adjuvant. The AE36 peptide was encapsulated in liposomes and its prophylactic and therapeutic effects were evaluated using mouse models. The liposomal formulations significantly increased the antitumor immune function of AE36 in both treatment and prophylaxis [56].
Alipour Talesh et al. [57] designed a cationic nanoliposome carrier system composed of 1,2-dioleoyl-3-trimethylammonium propane-cholesterol for vaccine formulation including natural multi-epitope HER2/neu-derived P5 peptide and poly-IC immunoadjuvant. The release of IFN-γ was higher in the vaccinated mouse group than in the control group. It was also observed that the nanovaccine formulation produced 2.7 times higher tumor growth delay in vaccinated mice.
A liposomal formulation composed of DMPC:DMPG: Chol:DOPE, encapsulated P5 and loaded MPL was used as a vaccine against BC on BALB/c mouse models. Enzyme-linked immunosorbent assay, cytotoxicity and intracellular cytokine assays, which followed immunization demonstrated that this formulation was unable to induce either CTL or CD4 T cell responses. Researchers concluded that using anionic conventional liposomes had resulted in a low uptake by DCs because of their negative surface charge [58].
Jalali et al. [59] evaluated the effecacy of four different multi-epitope HER2/neu peptide antigens (p5, p453, p373, and p1209) encapsulated in liposome-polycation-DNA (LPD) NPs for the induction of immune response in TUBO tumor mouse models. Two of the four tested peptides (p5 and p435)-LPD NPs caused higher IFN-γ release and cytotoxic responses in comparison with the control groups. Fifty percent of the mice that had been vaccinated with the p5-LPD and p435-LPD formulations remained tumor free for 80 days after eradiation of existing tumors. It was shown that 45% of NPs remained at the injection site after 24 hours, leading to a higher circulation time for the vaccine. The results of this study indicated that p5 and p435 peptides together with the nano delivery design had enhanced antigen-specific immunity.
Glaffig et al. [60] combined the tumor-associated MUC1 glycopeptide with the immunostimulating T-cell epitope P2 from tetanus toxoid, coupled to a dendrimer-like hyper-branched polyglycerol (hbPG) composed of azido-terminated triethyleneglycol acyl spacer and alkyne-functionalized hbPG-polymers. This structure led to improved water solubility of the vaccine formulation. A significant induction of immune response was observed in mouse models especially for the recognizing the human tumor cell (MCF-7) by IgG antibodies.
Nanodelivery systems are not only used in peptide antigen vaccines, but also in delivering gene-based vaccines. An example of such formulation is the lipid/calcium/phosphate NP-based mRNA vaccine encoding tumor antigen MUC1 combined with immune checkpoint blocker anti-CTLA-4 (CTL-associated protein 4) monoclonal antibody. Here, the researchers demonstrated that the nanovaccine formulation dramatically improved the anticancer immune response compared to the vaccine alone or monoclonal antibody alone, in TNBC mouse model [61].
Alginic acid-coated chitosan nanoparticles (A.C.NPs) have been used as oral delivery carriers for the legumain DNA vaccine to overcome its lack of efficiency in clinical application. Degradation of the vaccine in an acidic environment was evaluated in vitro and better protection of DNA vaccine via A.C.NPs has been proved by agarose gel electrophoresis. The antitumor effect of A.C.NPs-legumain DNA vaccine in BC mouse models was demonstrated by the lower tumor growth ratio and improved T lymphocyte responses [62].
Jadidi-Niaragh et al. [63], aiming to overcome the inefficacy of conventional DC-based BC vaccines caused by the immunosuppressive microenvironment of the tumor, used the vaccine in combination with CD73-specific small interfering RNA (siRNA)-loaded chitosan-lactate NPs. CD73-specific siRNA to decrease CD73 expression in tumor cells caused by its association with immunosuppression by adenosine, tumor-associated macrophages, regulatory T (Treg) and myeloid-derived suppressor cells. It augmented CTL responses, increased T cell proliferation and release of IFN-γ and IL-17, and enhanced the antitumor response. The results of this study indicated that 57% (4 of 7) of vaccinated mice showed significantly prolonged survival time with complete tumor remission.
Iranpour et al. [64] tested the ability of DCs, loaded with BC cell lysate in combination with three different poly-lactic-co-glycolic acid (PLGA) NP formulations, to induce antigen release, DC maturation, and T lymphocyte responses through these cells. It was shown that PLGA-NPs increased antigen presentation on DCs resulting in a higher amount of IFN-γ and IL-12 immunocytokine release from T lymphocytes. Kokate et al. [65] designed CpG-PLGA-NP-Tag (CpG-oligodeoxynucleotide [ODN]-coated tumor antigen [Tag] encapsulating NP) and evaluated the efficacy of this formulation ex vivo and in vivo. It was demonstrated that CpG-NP-Tag NPs were actively captured and presented by DCs. In vivo results showed a decrease not only in tumor size but also in angiogenesis, in addition to which an improved cytotoxic response was observed.
Unlike conventional approaches which use immunogen molecules in nanocarrier system, Campbell et al. [66] only encapsulated an endogenous protein derived from HMGB1 as an adjuvant in the NP. Hp91 peptide was packaged inside PLGA-NPs and its effectiveness in combination with HER2 antigen was evaluated. It was shown that Hp91-PLGA-NPs stimulated DCs and improved CTL and cytokine responses, compared to free HER2 peptide in vitro. In vivo results demonstrated that mice vaccinated with Hp91-PLGA-NPs and HER2 stayed tumor-free for a longer time, than mice vaccinated only with HER2.
Hartmann et al. [67] used cationic nanohydrogel particles to carry MUC1 antigen and CpG-ODN adjuvant in order to improve antitumor responses. They demonstrated that antigene loaded cationic nanohydrogel particles were immunologically neutral when used alone. By contrast, when the design NPs were loaded with MUC1 and CpG-ODN adjuvant, expression of co-stimulatory molecules and T cell proliferation were significantly boosted in vitro. Augmented antibody responses against MUC1 were also observed following in vivo vaccination of Balb/c mice.
Virus-like particles (VLPs) are some of the most intelligent nanotools with a great ability to encapsulate materials and chemically modify the external surface. VLP-based nanosystems are used for targeted delivery and controlled release of vaccine formulations in many types of infections and cancers [68]. Optimal nano-size, particulate nature and adjuvant activity of VLPs impart a great power to enhance strong immune responses to these particles [69].
Palladini et al. [70] developed a high-density display of HER2 antigene on the surface of VLPs and evaluated its prophylactic and therapeutic effects on mice. It was shown that the vaccine formulation overcomes B-cell tolerance and stimulates anti-HER2 IgG production. Fifty percent of the mice which were vaccinated with the HER2-VLP formulation survived without tumor growth for 1 year. The HER2-VLP vaccine has shown a promise in the prevention of tumor growth as also for the treatment of HER2+ BC.
Another approach using VLPs on BC immunotherapy was aimed at neutralizing relapse, metastatic progression and therapeutic resistance caused by BCSC. The cystine-glutamate antiporter protein xCT (SLC7A11) plays an important role for BCSC function. Bolli et al. [69] developed a vaccine formulation to target the xCT protein using a virus-like particle nanosystem (AX09-0M6), and proved its effectiveness in producing anti-xCT IgG2 antibodies and inhibiting tumor growth and pulmonary metastases (~42%) in mouse models.
Patel et al. [71] designed influenza VLPs modified to express BC HER2 converted to a glycosylphosphatidylinositol (GPI)-anchored form. A high level of T helper cell 1 and T helper cell 2 type anti-HER2 antibody response and protection against HER2-expressing tumor challenge in mouse models were because of GPIHER-2-VLP vaccination.
Go to :
CONCLUSION
Studies on immunotherapy have increased dramatically following the realization of the importance of cancer vaccines in overcoming the disadvantages of traditional therapies and provoking long-lasting protective antitumor immunity. Peptide-based vaccines, whole tumor cell vaccines, genetic-based vaccines and DC vaccines, are being evaluated for the use against BC, in both preclinical and clinical studies. Although the results of these vaccine studies show enhanced antitumor responses, it appears that some improvements may be needed owing to low immunogenicity of tumor antigens, genomic fusion risk of recombinant vaccines, absence of usable active adjuvants and the risk of uncontrolled vaccine release. It is known that even the weakest antigen, when used in combination with a suitable adjuvant, may produce stronger immune responses than the most immunogenic molecule acting alone. Therefore, development of new generation vaccine adjuvants may be essential. Additionally, each patient may have different BC phenotype because of the heterogenicity of this cancer. Accordingly, development of patient-specific vaccines may be very important. Immune tolerance and suppression pathways in the tumor microenvironment are also issues that need to be considered. Combining vaccines with other therapeutic agents as well as other antitumor immunity enhancing approaches may lead to better in-clinic responses. Furthermore, recent developments in nanothecnology show great promise as tools that may be used in developing effective immunotherapeutic vaccines for BC. Nanotechnology enables controlled release and targeted therapy, which are properties required to improve the efficacy of many types of BC vaccines. Efforts to develop an effective vaccine against BC may lead to improved antitumor clinical outcomes and therapeutic support for women fighting BC worldwide.
Go to :
 XML Download
XML Download