

 PDF
PDF Citation
Citation Print
Print



Abbreviations
C5
complement component 5
CM
conditioned media
HGSOC
high-grade serous ovarian cancer
ID8Luc
ID8 luciferase
KO
knockout
Mφ
macrophages
NIH
National Institutes of Health
OB
ob/ob
OC
ovarian cancer
OVX
ovariectomized
p53m
p53 mutant
TAMs
tumor-associated Mφ
TIMP1
tissue inhibitors of metalloproteinase-1
WT
wild-type
INTRODUCTION
The prevalence of obesity is rapidly and globally increasing (1). Obesity reduces human life expectancy and increases risks of multiple malignancies (23). In 2018, approximately 22,240 new cases and 14,070 deaths from ovarian cancer (OC) are expected in the United States (4). Despite OC being the 5th leading cause of cancer deaths among U.S. women and frequent spread through the peritoneal cavity rather than blood vessels (5), how obesity influences the progression of OC is not fully understood. Epidemiological evidence indicates controversial results between obesity and OC survival: both a positive relationship (6789) and no relationship (101112) have been reported. Despite this controversy, obesity appears as a risk factor for OC (131415) and leads to poorer quality-of-life outcomes in patients with OC (161718). Interestingly, obesity in early adulthood is linked with higher mortality in OC (121920). Obesity induces chronic inflammation which can lead to adverse health conditions such as cardiovascular disease, diabetes and some cancers. Obesity-associated inflammatory mediators may modify the tumor microenvironment, leading to cancer progression (2122). Adipose tissues release growth factors and cytokines such as adiponectin, leptin, TNF-α, IL-6 and IL-8/CXCL8 (23). Chronic inflammation has been shown to involve a chemokine network that influences the migration and invasion of cancer cells (2425), which support the tumor microenvironment for cancer progression by increasing inflammatory burden. We found that obesity could launch an inflammatory burden for OC progression via CXCR2-mediated signaling. OC cells produce high levels of CXCR2-specific chemokines such as CXCL1-3 and 8 (2627) in a NF-κB-dependent manner (2829). Furthermore, CXCR2-mediated signaling could be a central adipocyte-driven chemokine network (30), linking obesity and OC. Our recent study shows that CXCR2 is a negative regulator of p21 via Akt-mediated Mdm2 in OC, contributing to OC proliferation (31). High-grade serous ovarian cancer (HGSOC), the most common OC subtype, has highly mutated p53 tumor suppressor protein (up to 96%) (32). CXCR2-deriven progression of OC further enhances CXCR2-specific chemokines by potentiating NF-κB via epidermal growth factor receptor-transactivated Akt signaling (33). Functional restoration of p53 repressed CXCR2-specific chemokines CXCL1-3 and 8 in p53 mutant (p53m) OC cells (26). Our recent study indicates that CXCR2 associates with poor overall survival in p53m HGSOC (34), suggesting an involvement of CXCR2 in the high mortality of HGSOC compared to other OC subtypes. These facts indicate that obesity-induced chronic inflammation may enhance the inflammatory burden in OC followed by a high mortality via the accelerated progression of OC.
Because the relationship between obesity and OC survival remains controversial, we investigated if obesity could promote the progression of OC cells using the postmenopausal obese mouse model and the peritoneal dissemination of mouse ID8 OC cells, focusing on immune cell profile and cytokine signature in OC-induced ascites.
MATERIALS AND METHODS
Generation of stable ID8 luciferase (ID8Luc) OC cell line and culture
ID8Luc cells were generated from parental ID8 OC cells as described previously (35). Cells were cultured at 37°C in a water-saturated atmosphere of 95% air and 5% CO2 with DMEM containing penicillin/streptomycin (each 100 U/ml) and 4% FBS. All liquid culture media were acquired from Invitrogen (Grand Island, NY, USA).
Mouse peritoneal syngeneic model
Mouse peritoneal syngeneic model was performed under guidelines approved by the Institutional Animal Care and Use Committee at Meharry Medical College and the National Institutes of Health (NIH) guide for the Care and Use of Laboratory Animals. Wild-type (WT, C57BL/6J) and ob/ob mice (OB, B6.V-Lepob/J) were obtained from Jackson Laboratory (Bar Harbor, ME, USA). The mice were maintained in a specific pathogen free animal housing facility at 22°C±2°C and 40%–60% humidity under a 12:12 light: dark cycle. Nine-week old female mice were ovariectomized (OVX) for the peritoneal dissemination model of postmenopausal obesity and the OVX mice were maintained on a standard diet throughout the experimental period. ID8Luc OC cells (3×106 cells/mouse in a volume of 0.2 ml PBS) were injected intraperitoneally into WT and OB mice after complete recovery from OVX procedure and confirmation of weight gain. Bioluminescence imaging were monitored weekly for tumor growth. Briefly, mice anesthetized with 3% isoflurane were administered D-Luciferin (Cayman Chemical, Ann Arbor, MI, USA) intraperitoneally at 125 mg/kg, 5 min before acquisition of the image. Mice were then placed in the chamber of an In-Vivo MS FX PRO optical imaging system (Carestream, Rochester, NY, USA), and photons were collected for a period of 1 min. The luminescent intensity of the region of interest were quantified using Molecular Imaging software (Carestream). We monitored body weight and terminated the mice upon irreversible accumulation of ascites (up to 8–10 ml). Mice were monitored 3 times weekly to assess animal health such as hunched posture, lethargy and inactivity, impaired ambulation, shallow or labored breathing, hair coat condition and change in the body weight. In particular, mice showing clinical signs of ascites fluid production with constant increase of body weight and changes in appearance and activity were observed daily. When 20% increase in the body weight, extensive ascites accumulation or sluggish activity were observed, animals were terminated for humane reasons. Particularly, solid tumors from the diaphragm, omentum and pelvic sites were investigated for spreading index followed by histological examination using H&E staining. The survival time of the mice were compared between WT and OB mice with ID8Luc OC cells.
ELISA
CXCL1 levels in ascites and serum were measured by murine KC (CXCL1) ELISA kit (900-K127; PeproTech, Rocky Hill, NJ, USA) according to the manufacturer's instructions. The optical density of each well was determined using a microplate reader set to 405 nm with wavelength correction at 570 nm. Glucose levels in ascites and serum were measured by Glucose Colorimetric Assay Kit (Cayman Chemical). The optical density of each well was determined using a microplate reader at 514 nm wavelength.
FACS
Immune cell profiles in ascites were evaluated by FACS with the following specific antibodies for leukocyte subtypes: PerCP-Cy™ 5.5 Rat Anti-Mouse CD335 [NKp46] for NK cells; Alexa Fluor® 647 Rat Anti-Mouse F4/80 for macrophages (Mφ); PE Rat Anti-Mouse CD4 for CD4 T cells; and APC Rat Anti-Mouse CD8a for CD8 T cells (BD Biosciences, San Jose, CA, USA).
Proteomic array for cytokine signatures
Cytokine signatures in ascites were evaluated using Proteome Profiler Mouse Cytokine Array (ARY006; R&D Systems, Minneapolis, MN, USA).
PCR array and quantitative RT-PCR
PCR array for customized mouse chemokines (CAMP10242) was obtained from Qiagen (Frederick, MD, USA). After isolating total RNA from tumor samples and eliminating genomic DNA, the RT reaction was performed at 42°C for 15 min followed by 94°C for 5 min. A real-time PCR for chemokines was performed according to manufacturer's instructions using a Bio-Rad CFX96 (Hercules, CA, USA) and the following 2-step cycling program: 1 cycle at 95°C for 10 min, and 40 cycles at 95°C for 15 s and at 60°C for 1 min. Data analysis was performed based on a Data Analysis Center (https://www.qiagen.com/us/shop/genes-and-pathways/data-analysis-center-overview-page/) provided by Qiagen.
Adipogenesis
The mouse fibroblast cell line 3T3-L1 (CL-173) was purchased from the American Type Culture Collection. Cells were cultured in DMEM with 10% calf serum (Invitrogen) at 37°C in a water-saturated atmosphere of 95% air and 5% CO2, avoiding situations in which the cells became too confluent (>70%) before the initiation of differentiation. Differentiation was performed as described previously (30). Undifferentiated and differentiated cells were washed with sterile Dulbecco's phosphate-buffered saline and cultured for an additional 48 h in complete DMEM media followed by centrifuged conditioned media (CM) collection and storage at −80°C for future use.
Cell proliferation
Cell proliferation assay was performed using the cleavage of MTT to a colored product as described previously (31).
Cell migration
ID8 cells (2×105 cells/ml in serum-free DMEM with 1% BSA) to be used for a migration assay were seeded in the 24-well Transwell cell culture insert (VWR Corp., Radnor, PA, USA). The bottom chamber contained 0.5 ml DMEM supplemented with 10% FBS as a chemoattractant. Cells were treated as indicated in Results and then incubated for 24 h. The cells that remained inside the insert were removed with a cotton swab; migrating cells on the filter were fixed with 3.7% formaldehyde and stained with 0.1% crystal violet followed by washing of the cells with PBS. The number of migrating cells was counted under the microscope (×400) using 5 randomly chosen fields.
RESULTS AND DISCUSSION
Older women have higher rates of diagnosis and death from OC (36) and activation of estrogen receptor prevents fat accumulation (37). Because ovariectomy eliminates the protection of female mice from gaining body weight (38), we used OVX WT and OB mice on standard diets to reflect a postmenopausal status. In agreement with another study (39), obese mice gained 2-fold more body weight than WT mice (Fig. 1A). The OB mouse is leptin-deficient, resulting in obesity via hyperphagia despite a standard diet (40). We generated ID8Luc cells from parental mouse ovarian surface epithelial ID8 OC cells using stable transfection. Bioluminescence imaging revealed that tumors in obese mice were detected at earlier time points and were more widely spread metastasis of OC cells in the peritoneal cavity than lean mice (Fig. 1B). Bioluminescence intensity showed statistical significance in obese mice throughout the whole experimental period compared to lean mice (Fig. 1C). Other studies using the K18-gT121+/−; p53fl/fl; Brca1fl/fl (KpB) mice for serous epithelial OC model revealed that mice fed with a high-fat diet had larger tumor volume and heavier tumor weight than mice fed with a low-fat diet (4142), supporting our results showing more widespread metastasis of OC in the peritoneal cavity (Fig. 1B and C). Compared to lean mice (109 days), obese mice showed a shorter survival trend (97 days) following termination due to ascitic fluid accumulation as defined by humane endpoints (Fig. 2A). Ascitic fluid accumulation was easily detected in lean mice, while it was difficult to observe in obese mice due to a larger body size. This fact might have caused survival in obese mice to be overestimated. Obese mice had higher levels of glucose in both ascites and serum than lean mice (Fig. 2B), consistent with increased levels of blood glucose in mice fed with a high-fat diet (42). Obese mice had higher serum levels of CXCL1 but similar levels in ascites (Fig. 2C). Higher serum levels of CXCL1 indicate a systemic inflammatory burden in obese mice, which may promote cancer progression as suggested by our previous studies (263335). Similarly, increased serum CXCL1 and CXCL5 were associated with increased obesity in db/db mice (43). In addition, CXCL1 is required for obesity-dependent tumor adipose stromal cell recruitment, vascularization and tumor growth promotion (44). Obese mice had more tumor burden in the peritoneal cavity, showing the wider peritoneal dissemination of ID8Luc cells, compared to lean mice (Fig. 2D). These results are consistent with other report showing enhanced intraperitoneal tumor burden in overweight or obese mice (39). Furthermore, histological evaluation revealed that omental tumor tissues in obese mice had more adipocytes (Fig. 2D).
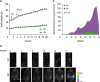 | Figure 1The peritoneal dissemination of OC in postmenopausal obese mouse model. (A) Body weight in WT (n=9) and OB (n=8) female mice. OVX WT and OB mice were fed with standard diets and generated mouse ID8Luc OC cells were injected intraperitoneally. (B) The tumor burden imaging in the peritoneal cavity of ID8Luc cell bearing mice measured by bioluminescence imaging. Representative pictures obtained from 9 WT and 8 OB mice. (C) Bioluminescence intensity in in the peritoneal cavity of ID8Luc cell bearing mice.
|
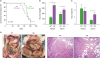 | Figure 2Parameters of the peritoneal dissemination of OC in postmenopausal obese mouse model. (A) Survival rate between WT (n=9) and OB (n=8) mice in the peritoneal dissemination model of OC cells. (B) Glucose levels in ascites and serum between WT and OB mice by ELISA. (C) CXCL1 levels in ascites and serum between WT and OB mice by ELISA. (D) Tumor burden and omental tumor tissues in the peritoneal cavity of ID8Luc cell bearing mice. Bold and thin yellow dots indicate the omental OC burden and the peritoneal OC dissemination, respectively. Representative pictures obtained from 9 WT and 8 OB mice.
*p<0.05.
|
We investigated immune cell profiles in ascites produced by OC. Ascites in obese mice showed higher levels of Mφ, but similar levels of NK, CD4 T and CD8 T cells compared to lean mice (Fig. 3). Multiple lines indicate that tumor-associated Mφ (TAMs) play an important role in progression of OC: TAMs constitute over 50% of cells in malignant ascites (45); CD163+ TAMs infiltration was associated with poor prognosis of OC (4647); high level of TAMs was associated with metastasis and advance of patients with OC (48); coculture of TAMs and SKOV3 OC cells increased the invasion ability of SKOV3 cells (48); M1-Mφ CM increased the metastatic potential, such as migration and invasion, in OC cells (49); and blocking Mφ function in mice with ID8 OC cells using a CSF-1 receptor kinase inhibitor (GW2580) reduced infiltration of M2-Mφ and decreased ascites volume (50). Based on these facts, obesity-promoted Mφ infiltration in ascites may accelerate OC progression followed by high mortality. Furthermore, we investigated chemokine and cytokine signatures in ascites from obese and lean mice. Ascites in obese mice expressed higher levels of CCL2, CXCL12, CXCL13, complement component 5 (C5), G-CSF, M-CSF, and tissue inhibitors of metalloproteinase-1 (TIMP1) compared to those in lean mice (Fig. 4). Obesity is known to promote breast cancer by CCL2-mediated Mφ recruitment (51). Mφ accumulation in adipose tissue induced by a high-fat diet were reduced in CCL2 knockout (KO) mice compared to WT mice (52). Circulating levels of CCL2 were elevated in human obese subjects (53). These results may support obesity-derived CCL2-mediated Mφ recruitment in ascites produced by OC.
 | Figure 3FACS analysis for immune cell profiles in the ascites of ID8Luc OC cell bearing WT (n=5) and OB (n=7) mice.
*p<0.05.
|
 | Figure 4Chemokine and cytokine signatures in the ascites of ID8Luc OC cell bearing WT (n=5) and OB (n=7) mice by proteomic analysis.
*p<0.05.
|
Diet-induced obese mice demonstrated a robust increase of CXCL12 expression in white adipose tissues, recruiting Mφ (54). CXCL12-induced adipose tissue Mφ chemotaxis was mediated by CXCR7 in obesity but not leanness (55). CXCL12-CXCR4/CXCR7 signaling axis might drive Myc-induced prostate cancer in obese mice (56). CXCL12 protein was found to be up-regulated during differentiation of THP-1 monocytes (57). Polarized CD163+ TAMs were associated with increased CXCL12 expression in gastric cancer (58). These results also may support obesity-derived CXCL12-mediated Mφ recruitment in ascites produced by OC. We found previously that the dominant chemokine in adipocytes was CXCL13 during adipogenesis in 3T3-L1 cells (30), being consistent with a high level of CXCL13 in mature adipocytes (59). Monocyte-like and mature Mφ were found to produce CXCL13 in inflammatory lesions (60). These facts suggest that CXCL13 in ascites of OC may be potentiated by obesity and Mφ accumulation. Although expression levels of C5 are similar in adipocytes from lean and obese subjects (61), obese dogs with obesity-related metabolic dysfunction show increased C5 protein levels (62). So far roles of C5 in obesity and OC are poorly understood.
Although Mφ may promote colon cancer growth via a GM-CSF/heparin-binding epidermal growth factor-like growth factor paracrine loop that is enhanced by CXCL12 (63), ascites from OC have low levels of GM-CSF (Fig. 4). G-CSF treatment reduced body weight and increased energy expenditure in a diabetic rat model (64). On the other hand, M-CSF KO mice showed reduced body weight and monocytes in blood and spleen but no change in neutrophils compared to WT mice (65). The small intestinal lamina propria of G-CSF receptor KO mice harbored reduced numbers of Mφ compared to those of WT mice, but levels of neutrophil-like cells were similar between these mice (66). These results indicate that G-CSF in ascites of OC may be potentiated by Mφ accumulation rather than obesity. In high-fat diet-fed mice, mature adipocyte-induced CCL2 and M-CSF increase M2-Mφ in melanoma tumors (67). M-CSF enhanced OC tumorigenesis and metastasis (68). High-fat feeding increased Mφ infiltration into adipose tissues and serum levels of M-CSF and TIMP1 in BALB/c mice (69). On the other hand, there was no change in the expression of M-CSF in the adipose tissue from obese mice and only a minor change in Mφ infiltration (70). Mφ recruitment was reduced in the uterus of osteopetrotic mutant mice lacking M-CSF (71). Elevated Mφ infiltration into omental fat was observable in lean women and exaggerated by obesity, being paralleled by CCL2 and M-CSF mRNA levels (72). These results support obesity-derived M-CSF-mediated Mφ recruitment in ascites produced by OC. TIMP1 KO mice with high-fat diet had a lower body weight and less subcutaneous and gonadal fat mass compared to WT mice (73). TIMP1 was highly secreted by omental adipose tissue in human obesity (74) and circulating levels of TIMP1 were higher in obese women than in lean women (75). These results indicate that TIMP1 in ascites of OC may be potentiated by obesity rather than Mφ accumulation.
We compared chemokine signatures and functional roles between preadipocyte and adipocyte CM-treated ID8 OC cells. A short-term treatment (1 h) of adipocyte-CM in ID8 OC cells showed above 5-fold induction in CCL7, CCL17, CCL20, CCL25, CCL27, CXCL1, CXCL2, CXCL3 and CXCL10 compared to preadipocyte-CM (Fig. 5A). Dominant chemokines (above 15-fold) induced by adipocyte-CM were CXCL2 and CXCL3 (Fig. 5A). Long-term treatment (24 h) with adipocyte-CM diminished chemokines induced by short-term treatment, although maintaining above 2-fold induction in CCL7 and CCL17 (Fig. 5A). High-fat diet fed mice bearing prostate tumors showed higher levels of CXCL1 and CXCL2 expression compared to low-fat fed mice (76). CXCL1 is required for the obesity-dependent tumor adipose stromal cell recruitment, vascularization and tumor growth promotion in prostate cancer (44). OC cell lines expressed highly CXCL1-3 and 8 (35), while ovarian tumor tissues expressed highly CCL20 (35) and CXCL10 (34). The differential chemokine signatures between adipocyte-CM on ID8 cells and ascites from obese mice with ID8 OC cells may be due to tumor heterogeneity including cancer cells, adipocytes, stromal cells and immune cells. Interestingly, adipocyte-CM prefers cell migration to viability compared to preadipocyte-CM in ID8 OC cells (Fig. 5A and B). Adipocyte-CM increased the proliferation and migration of ID8 cells compared to basal media not preadipocyte-CM (77). In B16F1 melanoma and E0771 breast cancer cells, adipocyte-CM supported cell proliferation and migration compared to basal media not preadipocyte-CM (78). In RM1 prostate cancer cells, adipocyte-CM increased cell proliferation but had no effect on migration compared to preadipocyte-CM (79). MCF7 and MDA-MB-231 breast cancer cells showed higher migration in a cancer-associated adipocyte-CM than in normal breast adipocyte-CM but no change in cell proliferation (80). In LNCaP prostate cancer cells, periprostatic explants CM enhanced cell motility but inhibited proliferation compared to stromal-vascular fraction CM (81). Despite cancer type-specific effects of adipocyte-CM on cancer cells, generally it looks like to enhance cell migration and somewhat proliferation in obesity-related cancer.
 | Figure 5Chemokine signatures and functional roles in preadipocyte- and adipocyte-CM treated ID8 OC cells. (A) Chemokine signatures in preadipocyte- and adipocyte-CM treated ID8 OC cells using PCR array containing complementary sequences for human chemokine genes. Expression levels of chemokines were defined as absent (light green), low (green area) and high (red area) on average threshold cycles. (B) Cell viability in preadipocyte- (PreA) and adipocyte-CM (Ad) treated ID8 OC cells n=3). (C) Cell migration in preadipocyte- (PreA) and adipocyte-CM (Ad) treated ID8 OC cells (n=3).
*Chemokines with >5 (1 h treatment) and >2 (24 h treatment)-fold increase were recognized as the major differences between preadipocyte- and adipocyte-CM; †p<0.05.
|
Based on our findings and other studies, obesity may accelerate the peritoneal dissemination of OC through systemically higher production of proinflammatory chemokines and increased Mφ recruitment in ascites.
 XML Download
XML Download