

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Dectin-1 is a C-type lectin receptor expressed on myeloid dendritic cells, macrophages/monocytes, T cells, and B cells. The Dectin-1 recognizes β-glucan of fungal cell wall particles, such as heat-killed Saccharomyces cerevisiae (HKSC), heat-killed Candida albicans, and zymosan, to protect fungal infection (12345678). Thus, recognizing β-glucan by Dectin-1 induces numerous cellular responses, including phagocytosis, respiratory burst, arachidonic acid metabolite production, and cytokine and chemokine induction, for promoting antifungal immunity. Antifungal antibodies are essential for the protection of hosts from pathogenic fungi (9101112).
Ig class switching is a process in which B cells shift from production of IgM to IgG3, IgG1, IgG2b, IgG2a, IgE, or IgA in mice or to IgG3, IgG1, IgA1, IgG2, IgG4, IgE, or IgA2 in humans (13). LPS, CD40 ligand, and various cytokines directly activate B cells and induce Ig class switching. In humans, IL-4 and IL-13 drive B cell switching to IgG4, IgE (141516), IgG3, and IgG1 (17), IL-10 to IgG3 and IgG1 (18), IL-13 to IgG4 and IgE (14), and TGF-β to IgA (1319). Moreover, IL-10, IL-12, IL-21, and vascular endothelial growth factor have been reported to skew class switching toward IgG4 (2021222324). This class switching is mediated by the class switch recombination (CSR) of the Ig heavy chain gene. The transcription of germline transcripts (GLT) on each switch region of the Ig heavy chain DNA in mature B cells is a prerequisite for each Ig CSR process (16). For instance, selective induction of GLTε transcription initiates IgE class switching by increasing the accessibility of activation-induced cytidine deaminase (AID), which is an essential enzyme for the Ig CSR process (25), to the non-transcribed DNA strand of the switch region.
We reported recently that Dectin-1 stimulation with its agonists (i.e., HKSC and depleted zymosan) selectively induces IgG1 class switching resulting in an increase of IgG1 production by mouse B cells (2627). Dectin-1 is also expressed on human B cells (2). However, the role of Dectin-1 in human B cells has not been determined. Here we found that direct Dectin-1 stimulation with the Dectin-1 agonist HKSC selectively induces GLTγ4 expression and IgG4 production by human B cells.
MATERIALS AND METHODS
Human B cell line and isolation of human tonsillar resting B cells
The mature human B cell line 2E2 (surface IgM+ and IgD+) (28) was provided by Dr. P. Casali (University of Texas Long School of Medicine, San Antonio, TX, USA). Fresh human tonsil tissues were obtained from tonsillectomies performed at the Department of Otorhinolaryngology–Head and Neck Surgery (Konyang University Hospital, Daejeon, Korea). The tonsil tissues were cut with sterilized scissors and homogenized by a homogenizer with HBSS (WelGENE, Daegu, Korea) containing 1% penicillin/streptomycin (Gibco, Invitrogen, Carlsbad, CA, USA). The suspended tonsillar cells were passed through a 70-µm cell strainer (BD Falcon, San Jose, CA, USA) to separate single cells. Tonsillar cells were prepared by a Ficoll/Histopaque-1077 (Sigma Aldrich, Saint Louis, MO, USA) density gradient method and further isolated by MACS negative selection using anti-CD43 microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany) to obtain untouched resting B cells. The purity of tonsillar resting B cells (CD43−CD19+, ≥98%) was assessed by flow cytometry using a FACSCalibur (BD Biosciences, San Jose, CA, USA), following staining of the cells with anti-human CD43 FITC (eBioscience, San Diego, CA, USA) and anti-human CD19 PE (BioLegend, San Diego, CA, USA). The Institutional Review Board of Konyang University Hospital approved this study (approval No. KYUH 2015-05-007-003).
Cell culture and reagents
Cells were cultured at 37°C in a humidified CO2 incubator (Forma Scientific, Marietta, OH, USA) in RPMI-1640 medium (WelGENE) supplemented with 10% fetal bovine serum (PAA Laboratories, Etobicoke, ON, Canada). Cells were stimulated using HKSC (1×107 cells/ml, InvivoGen, San Diego, CA, USA). Anti-human CD40 Ab was purchased from eBioscience and rhIL-4 was obtained from R&D Systems (Minneapolis, MN, USA). The Dectin-1 antagonist laminarin was purchased from InvivoGen.
RT-PCR
RNA preparation and RT-PCR were performed as previously described (27). PCR primers (Supplementary Table 1) were synthesized by Bioneer (Daejeon, Korea). The PCR for β-actin was simultaneously performed to normalize cDNA concentrations within each sample set. PCR products were resolved using electrophoresis on 2% agarose gels.
Reporter plasmid, transfection, and luciferase assays
The human GLTγ4 promoter DNA fragment (−1076 to +100) was amplified from human tonsil genomic DNA using PCR. PCR primers (Supplementary Table 1) were derived from previously reported human GLTγ4 promoter nucleotide sequences (2930). The GLTγ4 promoter fragment was subcloned into the pGL3-basic vector (Promega, Madison, WI, USA), and the reporter plasmid was named pGL3-hγ4[−1076/+100]. Transfection was performed by electroporation using a Gene Pulser II electroporation system (Bio-Rad, Hercules, CA, USA) as described previously (31). The reporter plasmid was co-transfected with pCMV-β-gal (Stratagene, La Jolla, CA, USA), and luciferase and β-gal assays were performed as described previously (31).
Isotype-specific ELISA
Abs produced in B cell cultures were detected using isotype-specific ELISAs as described previously (27).
Cell viability assay
Cell viability was determined using EZ-Cytox cell viability assay kits (Daeil Lab Service Co., Seoul, Korea) as described previously (32). Briefly, 20 µl of EZ-Cytox kit reagent was added to each cell-cultured well of a 96-well microplate and then incubated at 37°C in a humidified CO2 incubator for 3 h. After incubation, OD was measured at a wavelength of 450 nm using an absorbance microplate reader (BioTek Instruments, Inc., Winooski, VT, USA).
RESULTS AND DISCUSSION
Effect of HKSC on GLT expression in a human B cell line
First, we examined the effect of the Dectin-1 agonist HKSC on the expression of GLTs by the human B cell line 2E2. The 2E2 cells (IgD+IgM+) can undergo Ig CSR through initiation of GLT transcription after in vitro exposure to appropriate stimuli (2833). As shown in Fig. 1A, HKSC selectively induced GLTγ4 expression. AID mRNA expression was not affected by HKSC stimulation. Human Dectin-1 is alternatively spliced, resulting in several isoforms (splice variants: Dectin-1 [full length], Dectin-1b, and Dectin-1c) in peripheral blood mononuclear cells and immature monocyte-derived dendritic cells (343536). The expression of these isoforms is cell and activation specific, and Dectin-1b is mainly expressed in human dendritic cells and macrophages (2343637). The differences underlying specific isoform expression and function are unclear, although there are evidences for specific isoform-related functions in mice (53839). We observed that 2E2 B cells express Dectin-1b but not full-length Dectin-1 or Dectin-1c under basal conditions (Fig. 1A, lower left panel). Interestingly, Dectin-1c expression was dramatically induced by HKSC, and this expression is correlated with GLTγ4, whereas the basal expression of Dectin-1b was decreased. Hence, we speculate that HKSC-induced Dectin-1c mRNA expression has a certain critical role in inducing GLTγ4 transcription by human B cells. However, in a study with dendritic cells, Hermanz-Falcón et al. (34) described that Dectin-1c lacks a complete C-type lectin-like domain (CTLD); therefore, this spliced variant is unlikely to encode a functional lectin. The major function of Dectin-1 is likely carried out by Dectin-1 (full length) and Dectin-1b, both bearing a complete CTLD. Thus, the various Dectin-1 isoforms may serve specific roles in each immune cell type and differential isoform usage may represent a mechanism of regulating cellular responses to its ligand in the immune system (23538). Nonetheless, it will be important to address the function of the Dectin-1c isoform and the significance of the positive correlation between Dectin-1c and GLTγ4 expression in HKSC-stimulated B cells. On the other hand, the Dectin-1 antagonist laminarin abrogated HKSC-induced GLTγ4 and Dectin-1c expression (Fig. 1A, lower center panel). Next, we constructed a GLTγ4 promoter reporter and then determined whether HKSC activates the promoter activity. HKSC significantly enhanced GLTγ4 promoter activity (Fig. 1B). IL-4 treatment was used as a positive control for GLTγ4 induction (Fig. 1A, lower center panel and Fig. 1B). These results suggest that B cell Dectin-1 stimulation selectively induces GLTγ4 transcription through the regulation of Dectin-1b and Dectin-1c expression.
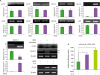 | Figure 1Dectin-1 agonist HKSC selectively induces GLTγ4 expression in human B cell line 2E2. (A) 2E2 cells were stimulated with HKSC (1×107 cells/ml), laminarin (10 µg/ml), and IL-4 (10 ng/ml). After 2 days of culture, mRNAs were isolated, and GLTs, AID, and Dectin-1 mRNA levels were measured by RT-PCR. Graphs indicate relative cDNA levels that are normalized to β-actin cDNA expression using ImageJ (National Institutes of Health, Bethesda, MD, USA) analysis. Densitometric data are averages of 2 independent experiments with ranges (bars). FL, full length. (B) 2E2 cells were transfected with the indicated human GLTγ4 promoter reporter (pGL3-hγ4[−1076/+100], 10 µg) and then stimulated with HKSC (1×107 cells/ml) and IL-4 (10 ng/ml). After 16 h, luciferase activities were analyzed. Data are presented as means±SEM of 3 independent transfections.
*p<0.05; **p<0.01.
|
Effect of HKSC on GLT expression and Ig production by primary human B cells
We used untouched tonsillar resting B cells (CD43−CD19+) purified from human tonsils to investigate the direct effect of the Decin-1 agonist HKSC on GLT expression and Ig production by primary human B cells. The purity of resting B cells assessed by flow cytometric analysis was higher than 98% (Fig. 2A). HKSC enhanced GLTγ4 and Dectin-1c expression by anti-CD40-stimulated B cells, and IL-4 also induced GLTγ4 (Fig. 2B). However, other GLTs were not induced by HKSC (data not shown). Furthermore, HKSC increased IgG4 production and decreased IgM and IgA production (Fig. 2C). Total IgG also tended to increase, but this seems to be due to an increase in cell viability by HKSC stimulation (Fig. 2C, lower right panel). These results indicate that HKSC selectively increases IgG4 production through the induction of GLTγ4 by human B cells. HKSC, a Dectin-1 agonist, may contain additional pathogen-associated molecular patterns that affect activation of other pattern recognition receptors of human B cells because it is a whole microorganism. Thus, the specific action of HKSC on Dectin-1 need to be further clarified.
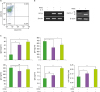 | Figure 2HKSC selectively enhances GLTγ4 expression and IgG4 production by human B cells. (A) The purity of isolated human tonsillar resting B cells (CD43−CD19+) was assessed using flow cytometric analysis. Resting B cells were stimulated with anti-CD40 Ab (5 µg/ml), HKSC (1×107 cells/ml), and IL-4 (10 ng/ml). After 4 days of culture, RNAs were isolated, and GLTγ4 and Dectin-1 mRNA levels were measured by RT-PCR (B). After 9.5 days of culture, supernatants were harvested, and Ig production levels were determined by isotype-specific ELISA (C). After 2.5 days of culture, cell viability (OD) was measured using the EZ-Cytox assay kit (C, lower right panel). Data are presented as means±SEM of 3 independent experiments.
*p<0.05; **p<0.01; ns: not significant.
|
IgG4 is the least abundantly found subclass of human IgG in normal serum. Many reports have demonstrated that IgG4 has unique structural and functional properties, such as anti-allergic (40414243444546) and anti-inflammatory (474849). IgG4 has the highest affinity for the inhibitory receptor FcγRIIB (48), which can have implications for inhibiting immune cells including B cells. Moreover, IgG4 plays a major role in the IgG Ab response in fungal infectious onychomycosis caused by Trichophyton (50). Thus, IgG4 can be used as a therapeutic Ab for allergic and inflammatory diseases and fungal infections (51525354). IgG4 is not clearly understood in human pathology, but elevated IgG4 levels are triggered in response to a chronic antigenic stimulus and inflammation. IgG4-related disease (IgG4-RD) is a chronic fibroinflammatory condition characterized by elevated serum IgG4 concentrations and tissue infiltration of IgG4-positive plasma cells that affects many organs (5556). IgG4-RD includes patients with autoimmune pancreatitis, hypophysitis, inflammatory aorticaneurysm, inflammatory pseudo-tumor, interstitial nephritis, interstitial pneumonitis, lymphadenopathy, Mikulicz's disease, prostatitis, retroperitoneal fibrosis, and Riedel thyroiditis. The immunopathogenesis of IgG4-RD has not been completely elucidated, and the role of IgG4 itself in disease pathogenesis remains unclear. A therapeutic agent used to treat IgG4-RD is rituximab, an anti-CD20 antibody, which depletes B cells. Rituximab therapy leads to rapid decline of serum IgG4 levels and prompt clinical improvement in IgG4-RD patients who do not respond to glucocorticoids, conventional steroidsparing agents, or both (57).
As previously mentioned, we have reported that HKSC selectively induces GLTγ1 transcription, IgG1 class switching, and IgG1 production by mouse B cells (2627). In the present study, we found that HKSC selectively induces GLTγ4 transcription and IgG4 production by human B cells. The functional characteristics of mouse IgG1 are very similar to human IgG4. Yet they, the ‘inactive’ isotypes, cannot activate complement by the classical pathway (i.e., they bind C1q very weakly and are also poor complement activators), bind more avidly to an inhibitory than to stimulatory FcRs, suppress immune complex deposition, and have limited ability to aggregate pathogens (48585960616263). Furthermore, we compared the sequences between human GLTγ4 and mouse GLTγ1 promoter. We found that there are highly conserved sequences (Supplementary Fig. 1). The nucleotide sequences of the 2 promoters showed an identity of 77.8%. The highly conserved sequences contain 3 previously identified NF-κB binding sites (293064) (underlined in Supplementary Fig. 1) and 2 putative C-Ets-1 binding sites. Together, these data raise the intriguing possibility that HKSC stimulation regulates NF-κB and C-Ets-1 signaling to activate transcription of both mouse GLTγ1 and human GLTγ4 by B cells. However, the underlying mechanisms need to be clarified.
In summary, our present study demonstrates for the first time the possibility that Dectin-1 can be involved in selective IgG4 class switching and IgG4 production by human B cells. Dectin-1 agonists including HKSC can be used as B cell adjuvants to augment IgG4 responses to control allergic, inflammatory, and fungal diseases. Moreover, a clear understanding of the mechanisms of Dectin-1 agonist HKSC-induced selective IgG4 production would contribute to the development of new therapeutic agents to modulate IgG4-RD.
 XML Download
XML Download