

 PDF
PDF Citation
Citation Print
Print



Abbreviations
AIM2
absent in melanoma 2
ALR
absent in melanoma 2-like receptor
ALS
amyotrophic lateral sclerosis
BMDM
bone marrow-derived macrophage
CARD
caspase recruitment domain
COPD
chronic obstructive pulmonary disease
DAMP
danger-associated molecular pattern
DSS
dextran sodium sulfate
EAE
experimental autoimmune encephalomyelitis
GSDMD
gasdermin D
IBD
inflammatory bowel disease
KO
knock-out
MOG
myelin oligodendrocyte glycoprotein
MS
multiple sclerosis
MSC
mesenchymal stromal cell
NLR
nucleotide-binding oligomerization domain-like receptor
OLG
oligodendrocyte
OMV
outer membrane vesicle
oxPAPC
oxidized phospholipids
PAMP
pathogen-associated molecular pattern
PD
Parkinson's disease
pro-caspase
pro-cysteine-aspartic protease
PRR
pattern recognition receptor
RA
rheumatoid arthritis
SOD1
superoxide dismutase 1
TRIF
TIR-domain-containing adaptor-inducing interferon-β
INTRODUCTION
Inflammation is an innate immune response that protects the body from invading pathogens and intracellular stress signals. It is characterized by redness, heat, swelling, pain, and loss of function (123). Inflammatory responses are initiated with the recognition of pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs) by pattern recognition receptors (PRRs) expressed on or inside innate immune cells (1345). Several types of PRRs, such as TLRs, have been found to express on cell surfaces and induce inflammatory responses through the recognition of their extracellular ligands, leading to the activation of several intracellular signaling molecules, such as NF-κB, AP-1, and IFN regulatory factor pathways (6789). By contrast, some PRRs localized in the cytoplasm, such as nucleotide-binding oligomerization domain-like receptors (NLRs), RIG-I-like receptors, and absent in melanoma 2 (AIM2)-like receptors (ALRs), induce inflammatory responses by recognizing intracellular ligands (41011).
In response to intracellular ligands, NLRs and ALRs induce a unique inflammatory response. These PRRs assemble intracellular protein complexes, known as “inflammasomes,” leading to gasdermin D (GSDMD)-mediated pyroptosis and activation of inflammatory caspase-1, which in turn induce the maturation and secretion of the pro-inflammatory cytokines IL-1β and IL-18 (41213). Inflammasomes can be classified into 2 major groups: canonical and non-canonical inflammasomes (451213). Because they are regarded as critical regulators of inflammatory responses, several studies have investigated their role in the pathogenesis of various human inflammatory diseases. However, the role of recently identified non-canonical inflammasomes, such as mouse caspase-11 and human caspase-4/5, remains poorly understood.
The present review aims to introduce the structure and activation mechanism of inflammasomes and to summarize and discuss the recent progress in understanding the regulatory role of caspase-11 non-canonical inflammasomes in the pathogenesis of inflammatory diseases. Moreover, this review highlights how selective targeting of caspase-11 non-canonical inflammasome could drive the development of effective therapeutics to prevent and treat human infectious and inflammatory diseases.
CASPASE-11 NON-CANONICAL INFLAMMASOME: STRUCTURE AND ACTIVATION
As discussed earlier, inflammasomes are classified into canonical and non-canonical inflammasomes. Canonical inflammasomes include members of NLR family, such as NLRP1, NLRP3, and NLRC4 and AIM2, and interact with pro-caspase-1, an effector of the inflammasome activation, with or without the bipartite adaptor, ASC. NLRP1 inflammasome is assembled by the interaction of the NLRP1, ASC, and pro-caspase-1 in response to Bacillus anthracis lethal toxin (41314). The assembly of NLRP3 and AIM2 inflammasomes is also similar to that of the NLRP1 inflammasome in response to several extracellular and intracellular ligands, such as pathogen nucleic acid hybrids, bacterial pore-generating toxins, extracellular ATP, oxidized mtDNA, mtROS, monosodium urate, cholesterol, potassium efflux, silica, crystals, alum, asbestos, β-amyloids, and hyaluronan (413141516) and cytosolic microbial double-stranded DNA (41314171819), respectively. NLRC4 inflammasome, however, is assembled by the direct interaction between NLRC4 and pro-caspase-1 in response to bacterial components, such as bacterial needle subunits, flagellin, and type III secretion apparatus (41314202122).
Both NLR family and AIM2 inflammasome belong to the canonical inflammasome class. Recently, a new inflammasome class was identified unexpectedly, when Kayagaki et al. (23) observed that toxin-induced inflammasome activation was totally inhibited in macrophages isolated from 129S6 mice. This mouse strain expresses non-functional truncated forms of caspase-11, because of polymorphisms in the caspase-11 gene locus. Additionally, caspase-11 mutant mice were found to exhibit resistance to LPS-induced endotoxic shock, by inhibiting the secretion of pro-inflammatory cytokines (24). Although caspase-11 inflammasomes activate inflammatory responses similar to canonical inflammasomes, they have been found to present unique properties and were thus classified as “non-canonical inflammasomes” (525). Caspase-11 is composed of an N-terminal CARD, a p20, and a C-terminal p10, and it is activated in response to intracellular LPS derived from gram-negative bacteria, such as Escherichia coli, Legionella pneumophila, Salmonella typhimurium, Shigella flexneri, Citrobacter rodentium, and Burkholderia spp. (525). Gram-negative bacteria generate outer membrane vesicles (OMVs) that deliver bacterial LPS into the cytosol of host cells by endocytosis (26). Caspase-11 inflammasome assembles by the direct interaction of caspase-11 CARD with lipid A moiety of the intracellular LPS, thereby leading to caspase-11-LPS complex oligomerization. As discussed earlier, caspase-11 was first identified in mice. Recently, several studies have suggested that caspase-4 and -5 may be human counterparts of murine caspase-11, based on the fact that, similar to caspase-11, they interact directly with intracellular LPS activating immune cell inflammasomes during inflammatory responses in a TLR4-independent manner (51125272829303132).
Although canonical and non-canonical inflammasomes differ both in structure and in activation mechanism, once active they activate the downstream inflammatory effector molecule caspase-1, by the proteolytic cleavage of inactive pro-caspase-1 to active caspase-1. The active caspase-1 then processes the inactive pro-inflammatory cytokines, pro-IL-1β and pro-IL-18 to the active IL-1β and IL-18 (4511131429). Moreover, inflammasome activation induces the cleavage of intracellular GSDMD at the aspartic acid 276 residue to produce GSDMD N-terminal fragments. The GSDMD N-terminal fragments in turn form pores in cell membranes, measuring about 20 nm in diameter. This induces pyroptosis, an inflammatory form of cell death (4525333435363738394041). The GSDMD-mediated pore formation also allows the secretion of the mature active pro-inflammatory cytokines, IL-1β and IL-18 from the inflamed cells (4525333435), which contribute to host defense by potentiating the inflammatory properties of phagocytes and triggering Th1 and Th17 cell-induced adaptive immune responses (42). Interestingly, recent studies have demonstrated that the pyroptosis regulator GSDMD is cleaved by caspase-1, and that the GSDMD N-terminal fragments are required for the transport and secretion of IL-1β and IL-18 across intact membranes of macrophages, dendritic cells, and neutrophils in a cell lysis-independent manner (4344). One study in particular investigated the cross-talk between canonical and non-canonical inflammasomes in inflammatory conditions, demonstrating that caspase-11-dependent IL-1β secretion is mediated by the activation of an NLRP3 inflammasome through the reduction of intracellular potassium levels (45). Another study identified a novel caspase-11 ligand and reported that caspase-11 interacts with not only intracellular LPS but also oxidized phospholipids (oxPAPC) in dendritic cells, leading to the caspase-11-dependent IL-1β secretion, but only LPS induces pyroptosis (46). The activation of caspase-4/5/11 non-canonical inflammasomes discussed is schematically summarized in Fig. 1.
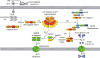 | Figure 1Activation of caspase-4/5/11 non-canonical inflammasomes. (A) LPS-harboring OMVs derived from gram-negative bacteria are endocytosed into the host cells, and (B) intracellular LPS directly interacts with mouse caspase-11 or human caspase-4/5 (caspase-4/5/11) through the caspase-4/5/11 CARDs and the lipid A moiety of intracellular LPS, (C) leading to the oligomerization of LPS-caspase-4/5/11 complexes to be activated. (D) Activated caspase 4/5/11 non-canonical inflammasomes cleave the linker loop of GSDMD at Asp276 residue to produce N- and C-terminal GSDMD fragments, and (E) the GSDMD N-terminal fragments generate GSDMD pores in the cell membranes by oligomerization, (F) resulting in cell swelling and rupture, known as pyroptosis. (G) Activated caspase 4/5/11 non-canonical inflammasomes also induce NLRP3 canonical inflammasome activation via an unknown mechanism, (H) leading to the proteolysis and activation of pro-caspase-1 to form active caspase-1 dimers. (I) Active caspase-1 dimer subsequently induces proteolysis and maturation of the inactive pro-inflammatory cytokines pro-IL-1β and pro-IL-18 to active IL-1β and IL-18, and (J) these active IL-1β and IL-18 are secreted through the GSDMD pores.N, GSDMD N-terminal fragment; C, GSDMD C-terminal fragment; PYD, pyrin domain; NACHT, nucleotide-binding and oligomerization domain; Asp276, aspartic acid 276 residue.
|
ROLES OF THE CASPASE-11 NON-CANONICAL INFLAMMASOME IN INFLAMMATORY DISEASES
As discussed earlier, inflammasome activation is critical for inflammatory response induction. Therefore, efforts have been made to treat inflammatory diseases by targeting inflammasomes, thus suppressing inflammatory responses. Most previous studies have focused on targeting canonical inflammasomes, in particular NLRP3. However, emerging studies have actively explored the role of recently identified non-canonical inflammasomes in inflammatory responses and evaluated them as promising targets to prevent and treat inflammatory diseases. This section will therefore discuss the recent studies that investigated the regulatory role of the caspase-11 non-canonical inflammasome in inflammatory diseases and its feasibility as a therapeutic target for these diseases.
Infectious diseases
Inflammation can be classified as sterile or non-sterile. Sterile inflammation is induced by cellular stress, injuries, and DAMPs, whereas non-sterile inflammation is induced by PAMPs originated from invading pathogens. This suggests that PAMPs derived from invading pathogens are critical to activate inflammatory responses. Infectious diseases are disorders caused by infection with various pathogens, such as bacteria, viruses, fungi, and parasites, which can trigger inflammasomes and their signaling pathways during inflammatory responses, thus inducing infectious disease pathogenesis. Several studies have explored the roles of caspase-11 in infectious diseases. Rathinam et al. (47) used caspase-11−/− mice to investigate the role of caspase-11 in infection by two gram-negative bacteria: E. coli and C. rodentium. They determined the molecular mechanisms underlying detrimental inflammatory responses, demonstrating that caspase-11 plays a critical role in the induction of inflammatory cell death and secretion of the pro-inflammatory cytokines IL-1α, IL-1β, IL-18, and high-mobility group box protein 1 by mouse bone marrow-derived macrophages (BMDMs) infected with gram-negative bacteria. Investigation of the molecular mechanisms revealed that through gram-negative bacterial infection, caspase-11 was activated by TIR-domain-containing adaptor-inducing interferon-β (TRIF)-mediated type I IFN signaling, subsequently synergizing with NLRP3 inflammasome assembly and activation, leading to caspase-1 activation (47). Interestingly, caspase-11-mediated inflammatory cell death was found to be caspase-1-independent (47). The study thus suggested that caspase-11 is critical to induce macrophage-mediated inflammatory responses during gram-negative bacterial infection, by coordinating with and activating TRIF signaling and canonical NLRP3 inflammasome. The same results were confirmed by Broz et al. (48), who generated caspase-11−/− and caspase-1−/− double knock-out (KO) mice and demonstrated the role of caspase-11 in macrophage-mediated inflammatory responses, as well as its underlying mechanism. The study reported that caspase-11 activation induced macrophage inflammatory death during S. typhimurium infection and that TRIF- and TLR4-dependent activation of IFN-β production is crucial to activate caspase-11 in macrophages. Moreover, caspase-11−/− and caspase-1−/− double KO mice were significantly less susceptible to S. typhimurium infection than caspase-1−/− KO mice (48), indicating that caspase-11-mediated inflammatory responses and cell death are detrimental for the host, irrespective of caspase-1-induced innate immune responses. Aachoui et al. (49) investigated the role of caspase-11 during infection with the cytosol-invasive bacteria Burkholderia thailandensis in caspase-11−/− and caspase-1−/− double KO mice. They looked into its underlying molecular mechanism, focusing on canonical inflammasomes. Their study reported that caspase-1 provided upstream priming signals to activate caspase-11, but not caspase-4 (i.e., the human ortholog of caspase-11) by inducing IL-18 and IFN-γ to clear B. thailandensis infection. Additionally, they showed that the caspase-11 activation is independent on the activation of canonical inflammasomes, such as NLRP3, NLRC4, and NLRC5. For caspase-11 non-canonical inflammasome-activated responses, the LPS of gram-negative bacteria needs to enter the cytosol of host cells by endocytosis and to be released from the endocytosed vacuoles. Aachoui et al. (49) investigated the differential activation of caspase-11 against gram-negative bacteria residing in the cytosol and vacuoles of mice infected with various gram-negative bacteria. The Burkholderia species of gram-negative bacteria naturally invades the cytosol (50), whereas several other gram-negative bacteria, such as S. typhimurium and L. pneumophilia, reside in the cytosolic vacuoles after infection (5152). Infection by gram-negative bacteria mutated to reside in the cytosol, such as S. typhimurium ΔsifA and L. pneumophilia ΔflaA ΔsdhA, triggered caspase-11 activation and caspase-11-mediated pyroptosis of mouse BMDMs and enhanced the clearance of infected bacteria independently of canonical inflammasomes, such as NLRP3 and NLRC4. Wild-type S. typhimurium infection, by contrast, caused a lower activation of caspase-11 than mutant S. typhimurium ΔsifA (53). Moreover, infection by the Burkholderia species also activated caspase-11, clearing the bacteria and protecting against bacterial septic shock in vivo (53). This indicates that caspase-11 is critical to sense cytosolic rather than vacuolar gram-negative bacteria. Taken together, these results suggest that caspase-11 is critical for the regulation of inflammatory responses, thus protecting the host against pathogen invasion.
Neurodegenerative diseases
Neurodegenerative diseases are a heterogeneous group of disorders caused by the progressive degeneration of the structure and function of the central and/or peripheral nervous system. They lead to cognitive and motor dysfunction and include multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Alzheimer's disease, and Parkinson's disease (PD). Neurodegenerative diseases share chronic inflammatory responses as a common cause, which are mediated by microglia — the resident macrophages of the nervous system. Activation of chronic inflammatory responses triggers neurotoxic signaling pathways that lead to a progressive degradation of the nervous system (54). Several studies have demonstrated that caspase-11 regulates the pathogenesis of neurodegenerative diseases. MS is an inflammatory demyelinating disease, which damages the protective insulating covers of central and peripheral neurons. Hisahara et al. (55) investigated the role of caspase-11 in the pathogenesis of MS, using mouse models of experimental autoimmune encephalomyelitis (EAE). EAE was induced in mice through immunization with the myelin oligodendrocyte (OLG) glycoprotein (MOG)35–55 peptide, and caspase-11 was highly expressed in OLGs, which are the myelinating cells of the central nervous system of EAE mice. Caspase-11 induced cytotoxic cytokine-mediated OLG cell death, to which the OLGs of caspase-11−/− mice were highly resistant (55). Moreover, susceptibility to EAE and inflammatory cytokine production by the central nervous system were markedly reduced in caspase-11−/− mice (55). These results indicate that caspase-11 activation induces demyelination in the central nervous system through OLS cell death facilitation, leading to MS pathogenesis. ALS, also known as Lou Gehrig disease or motor neuron disease, is an inflammatory neurodegenerative disorder leading to the death of the neurons responsible for voluntary muscle control. Kang et al. (56) investigated the role of caspase-11 in the pathogenesis of ALS using a mouse model. Superoxide dismutase 1 (SOD1) G93A transgenic mice were used as ALS models, and the inflammatory responses and disease symptoms were compared between wild-type and caspase-11−/− SOD1 G93A transgenic mice. Caspase-11−/− SOD1 G93A transgenic mice exhibited a lower activity of caspase-1 and caspase-3, as well as a lower level of IL-1β compared to control mice. Interestingly, inflammatory responses, neurodegeneration, and disease onset and progression did no differ significantly between caspase-11−/− and control SOD1 G93A transgenic mice (56), suggesting that, despite the contribution of caspase-11 for the pathology of ALS, its inhibition is not sufficient to prevent the disease. PD is a long-term inflammatory degenerative disorder of the central nervous system that affects the motor system, causing tremors, stiffness, slowness, impaired balance, and in advanced stages a shuffling gait. PD is known to be caused by the loss of dopamine-producing (dopaminergic) neurons in the substantia nigra of the midbrain. Furuya et al. (57) investigated the role of caspase-11 in the pathogenesis of PD using the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced neurotoxic mouse, which is an animal model of PD. They examined the function of caspase-11 on dopaminergic neuron death in caspase-11−/− diseased mice and found that those mice were more resistant to MPTP-induced neurotoxicity than wild-type mice. Moreover, microglial activation, production of the microglia-deleterious mediator IL-1β, and expression of the inflammatory gene inducible nitric oxide synthase were significantly attenuated in the substantia nigra of the MPTP-injected caspase-11−/− mice compared with MPTP-injected wild-type mice (57). This indicates that MPTP-induced neurotoxicity and dopaminergic neuronal cell death in the substantia nigra may be mediated by the activation of caspase-11 inflammatory signaling cascades.
Although the studies discussed above successfully demonstrated the critical roles of caspase-11 in the pathogenesis of neurodegenerative diseases MS, ALD, and PD, the mechanisms by which caspase-11 is potentially involved in the pathogenesis of these diseases is still unclear. There is no evidence in these studies to suggest a possibility of gram-negative bacterial infections or intracellular LPS activated the caspase-11 non-canonical inflammasome involved in the pathogenesis of these diseases. However, caspase-11 might be activated by naturally occurring intracellular molecules other than intracellular LPS in the inflammatory conditions, and these intracellular inflammatory molecules might bind directly with the caspase-11 and subsequently activate caspase-11 non-canonical inflammasome, leading to the pathogenesis of these neurodegenerative diseases. Indeed, despite the inhibitory action of oxPAPC on the caspase-11 non-canonical inflammasome activation, oxPAPC was found as a naturally occurring intracellular ligand directly binding with the caspase-11 (58), providing the possibility that there might exist direct caspase-11 ligands other than intracellular LPS to activate the caspase-11 non-canonical inflammasome in the pathogenesis of neurodegenerative diseases. Therefore, further studies to identify and validate the intracellular molecules directly binding with caspase-11 in the inflammatory conditions are necessary, and it will be important to demonstrate whether this binding could activate the caspase-11 non-canonical inflammasome, leading to the pathogenesis of the neurodegenerative diseases. Taken together, these results suggest that caspase-11 is activated during the pathogenesis of various inflammatory neurodegenerative diseases, and the selective blockade of caspase-11 could be an effective strategy to prevent and treat several (but not all) inflammatory neurodegenerative diseases.
Inflammatory bowel diseases
The group of inflammatory bowel diseases (IBDs) comprises inflammatory disorders of the colon and small intestine, and its typical types are ulcerative colitis and Crohn disease. The organs primarily affected by ulcerative colitis are the colon and the rectum, whereas those affected by Crohn disease are the small and large intestines (596061). IBD can be debilitating and may lead to life-threatening complications. The symptoms vary depending on the severity and location of inflammation and include diarrhea, fever, fatigue, abdominal pain and cramping, bloody stool, reduced appetite, and weight loss (596061). Recent studies have looked into the regulatory role of caspase-11 in IBD pathogenesis. Demon et al. (62) investigated the expression and function of caspase-11 in ulcerative colitis using dextran sodium sulfate (DSS)-induced colitis mice, a known animal model of colitis. Caspase-11 was constitutively expressed in the mice colon, and caspase-11−/− mice exhibited a considerably higher susceptibility to DSS-induced colitis than wild-type mice. Moreover, DSS-induced tissue damage and inflammatory cell infiltration were increased in the guts of caspase-11−/− mice, and the secretion of IL-1β and IL-18 was markedly reduced in both caspase-11−/− and caspase-1−/− colitis mouse models (62). These results suggest that caspase-11−/− mice are highly susceptible to colitis induction, and that inflammasome activation during colitis pathogenesis relies on the cooperation of caspase-11 and caspase-1. Williams et al. (63) used the same animal model to investigate the function of caspase-11 in gastrointestinal inflammation and colitis pathogenesis. DSS-induced colitis was significantly induced in the caspase-11−/− mice, moreover, the morbidity and colon inflammation were also increased in the caspase-11−/− mice compared to the control normal mice. Additionally, the levels of pro-inflammatory cytokines IL-1β and IL-18 were significantly reduced in the colon of caspase-11−/− mice, and research into the molecular mechanisms revealed that the hypersensitivity to colitis pathogenesis exhibited by these mice was strongly associated with the loss of these cytokines, and that the expression and function of caspase-11 contribute for disease attenuation (63). These results strongly suggest that caspase-11 is critical to protect the host from colonic inflammation and injury. Recently, Chen et al. (64) reported an interesting study investigating the regulatory roles of caspase-11 and NLRP3 inflammasomes in mesenchymal stromal cells (MSCs) during the pathogenesis of IBD. Both caspase-11 and NLRP3 inflammasomes were activated in bone-associated MSCs treated with inflammatory stimuli or isolated from bacteria-infected mice. However, only NLRP3 inflammasome was activated in bone-associated MSCs isolated from DSS-induced colitis mice, which showed an increased secretion of IL-1β and IL-18 and pyroptosis MSCs. Inhibition of these inflammasomes by the small molecule component 66PR in bone-associated MSCs improved the survival and therapeutic potential in DSS-induced colitis mice (64). These results indicate that MSCs play a pivotal role in the activation of inflammatory responses in an inflammasome-dependent manner, and that inflammasome inhibition in these cells may constitute a potential approach to treat IBD. Taken together, these studies suggest that caspase-11 non-canonical inflammasome plays a regulatory role in IBD pathogenesis, but more studies demonstrating the direct evidences that caspase-11 non-canonical inflammasome is a critical player in the pathogenesis of IBDs are highly demanded.
Rheumatoid arthritis (RA)
RA is a long-term inflammatory disease primarily affecting the joints of the knees, wrists, hands, and feet, and causing stiffness, swelling, and pain. RA is characterized by chronic synovial inflammation of the joints, leading to the destruction of cartilages and bones (6566). In some cases, RA can also damage parts of other body systems such as the eyes, lungs, heart, skin, and blood vessels (65). Although the causes and pathogenic mechanisms of RA have been extensively investigated, they remain poorly understood. However, pro-inflammatory cytokines, chemokines, inflammatory mediators, and matrix metalloproteinases are critically involved (6667), and efforts have been made to develop anti-RA therapeutics by targeting these inflammatory products. A recent study has demonstrated the regulatory role of caspase-11 in inflammatory responses during RA pathogenesis. Lacey et al. (68) investigated the role of caspase-11 and its downstream effectors on inflammatory responses and infectious condition in bacteria (B. melitensis)-induced inflammatory arthritis. Both caspase-11 and caspase-1 triggered joint inflammation and pro-inflammatory cytokine production in bacteria-infected inflammatory arthritis mice, and delayed joint inflammation was observed in bacteria-infected caspase-11−/− mice. Moreover, both caspase-11 and caspase-1 cleared the bacteria from the mice joints by activating IL-18 and inducing pyroptosis of joint macrophages (68). These results suggest that caspase-11 inflammasome induces joint inflammation in an IL-18-dependent manner, restricting the bacterial infection through the induction of macrophage pyroptosis, and that caspase-11 plays a pivotal role by actively involving in the inflammatory responses and pathogenesis of inflammatory arthritis.
Inflammatory respiratory diseases
Inflammatory respiratory diseases encompass the pathological conditions caused by inflammatory responses, which primarily affect the lungs and airways, such as the upper respiratory tract, trachea, bronchi, bronchioles, alveoli, and pleura. Inflammatory respiratory diseases range from mild to life-threatening conditions, such as common cold, bacterial pneumonia, idiopathic pulmonary fibrosis asthma, and chronic obstructive pulmonary disease (COPD) (697071). Several recent studies have reported the role of caspase-11 and the underlying molecular mechanisms in the pathogenesis of inflammatory respiratory diseases. Cheng et al. (72) investigated the role of caspase-11 in endotoxemia-induced inflammatory lung injury, using a mouse model of bacterial endotoxin (LPS)-induced acute lung injury. Endotoxin-mediated severe endothelial pyroptosis and inflammatory responses were found to be induced by the activation of caspase-11 and human caspase-4/5 inflammasomes in mice and human lung microvascular endothelial cells, and endotoxin-induced lung edema, neutrophil accumulation, and mortality were significantly reduced in the caspase-11−/− mice. In addition, bone marrow transplantation of wild-type hematopoietic cells in caspase-11−/− mice did not suppress endotoxemia-induced lung injury (72), indicating that endotoxemia is mediated by non-hematopoietic rather than hematopoietic caspase-11. Moreover, activation of caspase-11 induced mature IL-1β production and GSDMD cleavage in lung endothelial cells (72). These results demonstrate that the activation of the non-canonical inflammasomes caspase-11 and human caspase-4/5 in endothelial cells plays a central role to induce endothelial pyroptosis and tissue injury during the pathogenesis of endotoxemia-induced lung injury. Eltom et al. (73) investigated the role of caspase-11 in the pathogenesis of COPD, as well as the role of caspase-11 and its downstream effectors, caspase-1 and IL-1β/18 in cigarette smoke-induced airway inflammation using a cigarette smoke-exposed COPD mouse model. Cigarette smoke-induced secretion of IL-1α, IL-1β, and IL-18 and the airway neutrophilia, one of the hallmark symptoms of COPD (747576), were markedly attenuated in mice missing either caspase-11 or caspase-1/11 (73). Taken together, these studies suggest that caspase-11 and human caspase-4/5 inflammasomes are activated during the pathogenesis of inflammatory respiratory diseases, such as inflammatory lung injury and COPD. Therapeutic approaches targeting selectively the activation of non-canonical inflammasomes and their downstream effectors may comprise a promising strategy to treat inflammatory respiratory diseases.
CONCLUSION AND PERSPECTIVES
Inflammation includes a series of complex biological processes that detect and eliminate invading pathogens and cellular damage and stress; however, chronic inflammation, which is a slow and long-term process, is considered a major risk factor for several inflammatory diseases. Therefore, extensive efforts have been made to identify the molecular and cellular mechanisms by which inflammatory responses induce inflammatory diseases and to evaluate the molecular targets that modulate inflammatory responses, so as to develop safe and effective therapeutics for inflammatory diseases.
One critical event in inflammatory responses is the activation of inflammasomes. Recent studies have demonstrated that unlike canonical inflammasomes, caspase-4/5/11 non-canonical inflammasomes are activated by intracellular LPS. Similar with canonical inflammasomes, non-canonical inflammasomes induce caspase-1 activation and GSDMD cleavage, leading to GSDMD-mediated pyroptosis through the generation of cell membrane pores and caspase-1-mediated maturation and secretion of the pro-inflammatory cytokines, IL-1β and IL-18, suggesting that selective and effective targeting of the non-canonical inflammasomes and their downstream effectors, such as caspase-1, GSDMD, and pro-inflammatory cytokines, could be considered as a potential and promising approach to suppress inflammatory responses, thus treat inflammatory diseases.
Indeed, several studies have successfully demonstrated that targeting inflammasome signaling cascades effectively suppresses inflammatory responses and potentially ameliorates the symptoms of various inflammatory diseases, however, most studies have focused on canonical inflammasomes, especially NLRP3 inflammasome. Few studies have explored the regulatory roles of non-canonical inflammasomes in limited types of inflammatory conditions, such as infectious diseases, neurodegenerative diseases, IBDs, inflammatory arthritis, and inflammatory respiratory diseases, as summarized in Table 1. Therefore, further studies are needed to demonstrate the role of non-canonical inflammasomes in various inflammatory diseases, as well as the functional crosstalk between canonical and non-canonical inflammasome signaling cascades in these inflammatory conditions. Moreover, LPS has been reported as the only ligand to activate caspase-4/5/11 non-canonical inflammasomes in macrophages, and the studies identifying new molecules to activate the caspase-4/5/11 non-canonical inflammasomes and demonstrating how the activation of caspase-4/5/11 non-canonical inflammasomes by these ligands induce the pathogenesis of inflammatory diseases in different ways of canonical inflammasomes are highly required. In addition, most studies investigating the role of non-canonical inflammasomes in inflammatory diseases have been conducted in cell systems and animal disease models. Studies in human subjects and inflammatory disease patients have been very limited, with no current reports of the functions of non-canonical inflammasomes. This highlights the need for clinical trials evaluating the regulatory role of non-canonical a well as canonical inflammasomes. Finally, most previous studies demonstrated the role of non-canonical inflammasomes in inflammatory responses and diseases by deleting caspase-4/5/11 genes in mice. Therefore, the need to develop safe and effective therapeutics that selectively target and modulate the activities of non-canonical inflammasomes and their downstream effectors remains, to inhibit inflammatory responses in inflammatory diseases.
Table 1
Roles of the caspase-11 non-canonical inflammasome in infectious and inflammatory diseases

| Diseases | Models | Caspase-11 roles | Ref. |
|---|---|---|---|
| Infectious | E. coli or C. rodentium-infected caspase-11−/− mice | 1. Caspase-11 induced pyroptosis and pro-inflammatory cytokine secretion by gram-negative bacterial infection | (47) |
| 2. Caspase-11 induced assembly and activation of NLRP3 inflammasome to induce inflammatory responses | |||
| 3. Caspase-11 was activated by TRIF-induced type I IFN | |||
| 4. Caspase-11-induced pyroptosis was caspase-1-independent | |||
| S. typhimurium-infected caspase-11−/− mice | 1. Caspase-11 induced pyroptosis and pro-inflammatory cytokines by gram-negative bacterial infection | (48) | |
| 2. Caspase-11 increased the susceptibility to the S. typhimurium infection-induced inflammatory responses | |||
| 3. Caspase-11 was activated by TRIF- & TLR4-induced IFN-β | |||
| 4. Caspase-11-induced pyroptosis was caspase-1-independent | |||
| B. thailandensis-infected caspase-11−/− mice | 1. Caspase-11 expression was increased by IFN-γ produced by caspase-1-activated IL-18 by gram-negative bacterial infection | (49) | |
| 2. Caspase-11 activation was independent on the activation of canonical inflammasomes | |||
| Caspase-11−/− mice infected with either wild-type (Burkholderia species, S. typhimurium and L. pneumophilia) or mutant bacteria (S. typhimurium ΔsifA and L. pneumophilia ΔflaA ΔsdhA) | 1. Caspase-11 induced pyroptosis by gram-negative bacterial infection | (53) | |
| 2. Caspase-11 protected against bacterial septic shock in mice by clearing infected bacteria | |||
| Neurodegenerative | MOG35–55 peptide-induced EAE caspase-11−/− mice | 1. Caspase-11 expression was increased in EAE mice | (55) |
| 2. Caspase-11 induced cytotoxic cytokine-mediated OLG cell death | |||
| 3. Caspase-11 increased EAE susceptibility and inflammatory cytokine production | |||
| SOD1 G93A transgenic caspase-11−/− mice | 1. Caspase-11 induced caspase-1 activation and IL-1β secretion in ALS mice | (56) | |
| 2. Caspase-11 did not induce neurodegeneration and ALS onset/progression | |||
| MPTP-injected neurotoxic caspase-11−/− mice | 1. Caspase-11 induced the susceptibility to the MPTP-induced neurotoxicity in mice | (57) | |
| 2. Caspase-11 induced microglial activation, IL-1β production, and iNOS expression in the substantia nigra of the MPTP-induced neurotoxic mice | |||
| IBD | DSS-induced colitis caspase-11−/− mice | 1. Caspase-11 was constitutively expressed in the colitis mice | (62) |
| 2. Caspase-11 lowered the susceptibility to colitis | |||
| 3. Caspase-11 increased the tissue damage and inflammatory cell infiltration in the guts of the colitis mice | |||
| 4. Caspase-11 reduced the secretion of IL-1β and IL-18 in the colitis mice | |||
| DSS-induced colitis caspase-11−/− mice | 1. Caspase-11 inhibited colitis pathogenesis in mice | (63) | |
| 2. Caspase-11 decreased the morbidity and the colon inflammation in the colitis mice | |||
| 3. Caspase-11 induced the production of IL-1β and IL-18 in the colons of the colitis mice | |||
| 1. Bone-associated MSCs | 1. Caspase-11 was not activated in the bone-associated MSCs of the colitis mice | (64) | |
| 2. DSS-induced colitis caspase-11−/− mice | |||
| RA | B. melitensis-infected inflammatory arthritic caspase-11−/− mice | 1. Caspase-11 induced the joint inflammation and the pro-inflammatory cytokine production in the bacteria-infected inflammatory arthritic mice | (68) |
| 2. Caspase-11 cleared the bacteria from the joints of the inflammatory arthritic mice by inducing IL-18 production and pyroptosis | |||
| Respiratory | hMVECs | 1. Caspase-11 induced the endothelial pyroptosis and inflammatory responses in the lung injury mice and the hMVECs | (72) |
| LPS-injected lung injury caspase-11−/− mice | 2. Caspase-11 induced the lung edema, the neutrophil accumulation, and the mortality in the lung injury mice | ||
| 3. Caspase-11 induced the IL-1β production and the GSDMD cleavage in the mouse lung endothelial cells | |||
| Cigarette smoke-exposed caspase-11−/− mice | 1. Caspase-11 induced the secretion of IL-1α, IL-1β, and IL-18 in the COPD mice | (73) | |
| 2. Caspase-11 induced the airway neutrophilia in the COPD mice |
![]()
In conclusion, given the existing evidence on the regulatory roles of non-canonical inflammasomes during inflammatory responses, selective targeting of non-canonical inflammasomes by novel pharmacological agents may potentially be used to prevent and treat various human inflammatory diseases.
 XML Download
XML Download