

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Inflammatory bowel disease (IBD) is an autoimmune disease resulting from chronic and inappropriate inflammatory response to intestinal microbes affected by environmental factors and genetic susceptibility of host.123 Forkhead box P3 (FOXP3+) regulatory T (Treg) cells and T helper 17 (Th17) cells possess anti-inflammatory and pro-inflammatory effects, respectively, in autoimmune diseases.14 It has been suggested that the loss of balance between these regulatory and effector T cells can lead to the development of Crohn's disease and ulcerative colitis.45
It is known that FOXP3+ Treg cells are increased in inflamed mucosa of IBD.67 Gil et al.8 have reported that FOXP3+ Treg cells are increased in the lamina propria (LP) and lymphoid aggregates of follicles (LF) in the lesion of Crohn's disease. Although FOXP3+ Treg cells are increased in the lesion of IBD, they have normal regulatory functions. Why these cells fail to regulate inflammation in IBD is currently unknown.9 Th17 cells might also be increased in inflamed mucosa of IBD.10
Cytokines play crucial roles in the pathogenesis of IBD.11 Various cytokines produced by innate immune cells and T cells have pro-inflammatory or anti-inflammatory functions.1112 Cytokines made by innate immune cells can recruit naïve CD4+ T cells and let them differentiate into effector T cells, such as Th1, Th2, Th17, FOXP3+ Treg, and follicular helper T (Tfh) cells.11 Cytokines excreted by differentiating naïve CD4+ T cells after antigen presenting to innate immune cells can stabilize and polarize themselves into specified effector T cells.11 Finally, effector T cells will make signature cytokines that are mediators with specified roles.11 The objective of this study was to investigate mucosal immunity related to Treg cells and Th17 cells and perform cytokine analysis in pediatric IBD.
METHODS
Among 37 candidate children who visited Jeju National University Hospital from April 2015 to December 2017, 12 children with IBD (9 with Crohn's disease and 3 with ulcerative colitis) and 7 healthy children as controls were enrolled. Their peripheral blood samples were collected. Serum was separated from the sample and stored at −80.0°C. Mucosal tissue was collected and soaked in 4% paraformaldehyde solution. All patients were on the stage of initial diagnosis or aggravation of IBD. Exclusion criteria were patients with mucosal healing state of IBD or any other underlying disease such as chronic granulomatous disease and bacterial enterocolitis. Their mean age (mean ± standard deviation [SD]) was 12.7 ± 0.8 years old in the control group, 14.9 ± 1.5 years old in the Crohn's disease group, and 15.8 ± 0.7 years old in ulcerative colitis group (Table 1).
Table 1
Demographic characteristics of patients with IBD and control subjects

IBD = inflammatory bowel disease, PCDAI = pediatric Crohn's disease activity index, PUCAI = pediatric ulcerative colitis activity index, CRP = carbohydrate reactive protein, ANCA = anti-neutrophil cytoplasmic antibody, ASCA = anti-saccharomyces cerevisiae antibody, ANA = anti-nuclear antibody, Dx. = diagnosis, FGIDs = functional gastrointestinal diseases including irritable bowel syndrome and functional abdominal pain.
![]()
Immunohistochemical staining
Paraffin-embedded sections were deparaffinized and dehydrated. Antigen retrieval was then performed in citrate buffer (pH 6.0) for 20 minutes using a microwave. Next, sections were incubated in 0.3% hydrogen peroxide and horse serum of Vector Elite ABC Kit (Vector Laboratories, Burlingame, CA, USA) to block endogenous peroxidase and unspecific responses. Anti-human FOXP3 (mouse monoclonal antibody, dilution 1:200; Abcam, Cambridge, UK) or anti-RORγ (rabbit polyclonal antibody, dilution 1:100; GeneTex Inc., CA, USA) was used as primary antibody.
After incubation with the primary antibody at 4.0°C for 16–18 hours (overnight), sections were treated with biotinylated anti-mouse or rabbit IgG antibody of Vector Elite ABC Kit (Vector Laboratories) at room temperature for 45 minutes. After washing with phosphate-buffered saline (PBS) containing 0.3% Triton X-100 (Junsei Chemical, Tokyo, Japan), slides were incubated with streptoavidin conjugated peroxidase of the same kit for 45 minutes at room temperature. Then 3,3′-diaminobenzidine (DAB, Dako, Glostrup, Denmark) was used as a substrate for color development. After color development, these sections were counterstained with Mayer's hematoxylin (Sigma-Aldrich, St. Louis, MO, USA) and mounted in Canada balsam (Junsei Chemical). Three lesions showing the most representative staining from each section were analyzed under higher magnification to count positive cells using a light microscope (LeicaDM LB2; Leica, Wetzler, Germany) and Image J software (National Institute of Health, Bethesda, MD, USA).
Cytokine analysis with multiplex assay
A multiplex kit (Bio-Plex Human Th17 cytokine panel 15 plex; Bio-Rad, CA, USA) was used to analyze cytokine polarizing Th17 cells and FOXP3+ Treg cells. This 15-plex panel could analyze 15 cytokines: interleukin (IL)-1β, IL-4, IL-6, IL-10, IL-17A/F, IL-21, IL-22, IL-23, IL-25, IL-31, IL-33, interferon (IFN)-γ, sCD40L, and tumor necrosis factor (TNF)-α. Serum preparation was done one day before the multiplex assay. Serum samples of patients and control that were stored at −80.0°C were thawed at room temperature and then centrifuged at 15,000 rpm for 15 minutes at 4.0°C. Then 100 µL of supernatant from the each sample was transferred to a 96-well plate and stored at 4.0°C. Laboratory procedures were then performed following the manufacturer's instructions. After making pre-mixed standard (S) from S1 to S8 with one fourth dilution each 7 steps, Th17 15-plex magnet beads (1X) from 20X was prepared by dilution using assay buffer. The standard and serum samples were stabilized at the room temperature for 20 minutes. Then 50 µL of 15-plex beads (1X) was transferred to wells. Two times of washing with 100 µL of wash buffer were done by adding buffer to wells. After placing the 96-well plate to a magnet holder, the buffer was taken out. After transferring 50 µL of standard and samples to the wells of the plate, the plate was sealed with sealing tape and wrapping with an aluminum foil followed by incubation on a shaker (1,100 rpm) at room temperature for 60 minutes.
After peeling off the sealing tape, three times of washing (100 µL of wash buffer each) were done. Then 25 µL of detection antibody (1X) was transferred to each well. After sealing the plate with sealing tape and wrapping it with aluminum foil, the plate was placed on a shaker (1,100 rpm) at room temperature for 30 minutes. After peeling off the sealing tape, three times of washing (100 µL of wash buffer each time) were done. After vigorous vortexing streptavidin-PE (1X), 50 µL of the solution was added to each well. After sealing the plate with sealing tape and wrapping it with aluminum foil, incubation was done on a shaker (1,100 rpm) at room temperature for 10 minutes. After peeling off the sealing tape, three times of washing with 100 µL of wash buffer each time was done. Then 125 µL of assay buffer was added to each well and the plate was covered with sealing tape. The plate was placed on a shaker (1,100 rpm) at room temperature for 30 seconds. Finally, the plate was transferred to a Bio-Plex Reader (Bio-Rad, CA, USA) for reading.
Statistical analyses
Statistical analysis was done using SPSS version 18.0 (SPSS Inc., Chicago, IL, USA). Results are presented as mean ± standard error or mean ± SD for each group. Statistical comparison between two groups and three groups was done using Student's t-test and Kruskall-Wallis test, respectively. Statistical significance was considered at P < 0.05.
RESULTS
Immunohistochemical staining
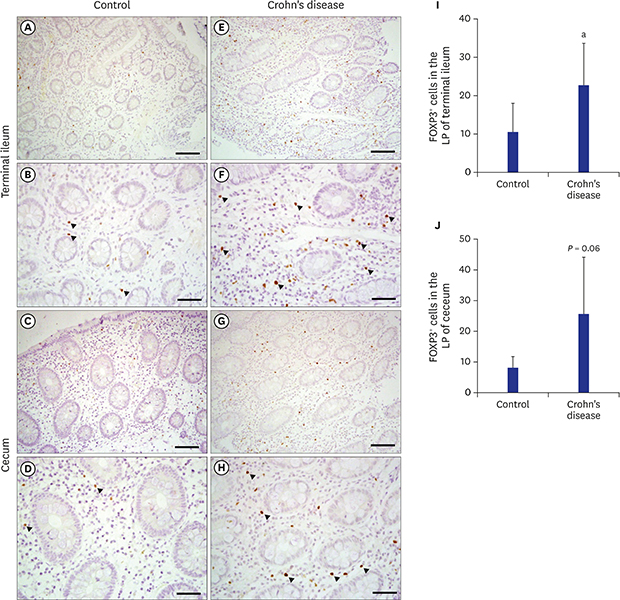
In immunohistochemical staining for FOXP3+ Treg cells in the LP of the terminal ileum (P < 0.05) and cecum (P = 0.06), the frequency of Treg cells was significantly higher in the terminal ileum of Crohn's disease group than that in the control (Fig. 1). In immunohistochemical staining of FOXP3+ Treg cells for LF of the terminal ileum, there was no significant difference between Crohn's disease group and the control (Fig. 2). Although the frequency of RORγt+ T cells polarizing Th17 cells in the terminal ileum of Crohn's disease group tended to be higher than that in the control, the difference between the two was not statistically significant (Fig. 3).
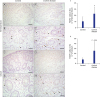 | Fig. 1Immunohistochemical staining of FOXP3 in the LP of terminal ileum and cecum in the control group (A-D) and the Crohn's disease group (E-H). Scale bars = 100 µm (A, C, E, G) and 50 µm (B, D, F, H). (I, J) Quantification of FOXP3 positive cells performed using three lesions showing the most representative expression of each patient in (I) terminal ileum and (J) cecum. Data are represented as means ± standard error.FOXP3+ = forkhead box P3, LP = lamina propria.
aP < 0.05 compared to the control group by student's t-test.
|
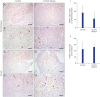 | Fig. 2Immunohistochemical staining of FOXP3 in the LF of terminal ileum and cecum in the control group (A-D) and the Crohn's disease group (E-H). Scale bars = 100 µm (A, C, E, G) and 50 µm (B, D, F, H). (I, J) Quantification of FOXP3 positive cells performed using three lesions showing the most representative expression of each patient in (I) terminal ileum and (J) cecum. Data are represented as means ± standard error.FOXP3+ = forkhead box P3, LF = lymphoid aggregates or follicles.
|
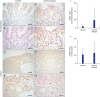 | Fig. 3Immunohistochemical staining of RORγ in the terminal ileum and cecum in the control group (A-D) and the Crohn's disease group (E-H). Scale bars = 100 µm (A, C, E, G) and 50 µm (B, D, F, H). (I, J) Quantification of RORγ positive cells performed using three lesions showing the most representative expression of each patient in (I) terminal ileum and (J) cecum. Data are represented as means ± standard error.
|
Cytokine analysis with multiplex assay
Results of multiplex assay are shown as Fig. 4. The results revealed that serum concentrations of IL-4 (9.6 ± 1.5 vs. 12.7 ± 3.0 pg/mL; P = 0.013), IL-21 (14.9 ± 1.5 vs. 26.4 ± 9.1 pg/mL; P = 0.006), IL-33 (14.3 ± 0.9 vs. 19.1 ± 5.3 pg/mL; P = 0.002), and IFN-γ (15.2 ± 5.9 vs. 50.2 ± 42.4 pg/mL; P = 0.002) in Crohn's disease were significantly lower than those in the control (Fig. 4A, D, F, and G). However, serum concentration of IL-6 (119.1 ± 79.6 vs. 52.9 ± 39.1 pg/mL; P = 0.030) in Crohn's disease was higher than that in the control (Fig. 4B). Serum concentrations of IL-17A (64.2 ± 17.2 vs. 28.3 ± 10.0 pg/mL; P = 0.012) and IL-22 (37.5 ± 8.8 vs. 27.2 ± 3.7 pg/mL; P = 0.022) in ulcerative colitis were significantly higher than those in Crohn's disease (Fig. 4C and E) while serum concentration of IL-33 (13.8 ± 0.28 vs. 19.1 ± 5.3 pg/mL; P = 0.015) in ulcerative colitis was significantly lower than that in the control (Fig. 4E). The serum TNF-α concentration was not different between the control and IBD groups (control group 21.5 ± 11.1 pg/mL; Crohn's disease group 21.6 ± 13.7 pg/mL; and ulcerative colitis group 16.3 ± 6.8 pg/mL; P = 0.917).
 | Fig. 4The cytokine analysis with multiplex assay shows the difference of cytokine concentration among the control, Crohn's disease, and UC. The serum concentration (pg/mL) of IL-4, IL-21, IL-33, and IFN-γ were significantly lower in Crohn's disease than the control (A, D, F, G). The serum concentration of IL-6, however, was higher in Crohn's disease than the control (B). The serum concentrations of IL-17A and IL-22 were significantly higher in the UC than those in Crohn's disease (C, E). The serum concentration of IL-33 was significantly lower in the UC than the control (E). Kruskall-Wallis test and Mann-Whitney test for post hoc analysis, statistically significant: P < 0.05.UC = ulcerative colitis, IL = interleukin, IFN = interferon.
|
DISCUSSION
In our study, the frequency of FOXP3+ Treg cells of LP in the terminal ileum of Crohn's disease was significantly higher than that in the control (Fig. 1). A few studies have reported that the number of FOXP3+ Treg cells in inflamed mucosa of IBD is increased based on immunohistochemical staining.67813 This paradoxical increase of FOXP3+ Treg cells suggests functional deficits of these cells. Some authors have reported dysfunction of these cells in IBD.14 However, other authors have reported that these cells have normal regulatory functions in vitro.151617 Although the therapeutic efficacy of Treg cells in IBD has been revealed recently,1819 the role of Treg in IBD remains controversial.9
The frequency of RORγt+ T cells that can differentiate to Th17 cell tended to be higher in inflamed mucosa of Crohn's disease than that in the control in the present study based on immunohistochemical staining (Fig. 3). However, the difference between the two was not statistically significant. Although a few reports have suggested that Th17 cell infiltration in inflamed mucosa of IBD is increased, the result is based on polymerase chain reaction of mRNA of Th17 cells from inflamed mucosa, not based on immunohistochemical staining.410
Cytokines play crucial roles in the pathogenesis of IBD.11 Results of serum cytokine analysis with the multiplex assay revealed that serum concentrations of IL-4, IL-21, IL-33, and IFN-γ in Crohn's disease were significantly lower than those in the control (Fig. 4A, D, F, and G). IL-4 and IL-33 are polarizing cytokines for the differentiating of naïve CD4+ T cells into Th2.2021 Our results are similar to those of previous reports.2223 West et al.23 have revealed that serum IL-4 concentration is decreased in the LP mononuclear cells of inflamed tissue of IBD. Matsuure et al.22 have reported selective resistance of mucosal T-cell activation to immunosuppression of IL-4 in Crohn's disease, but not in the control or ulcerative colitis. IFN-γ is a polarizing cytokine for differentiating of naïve CD4+ T cell into Th1.124 IFN-γ, a pro-inflammatory cytokine playing a role in the pathogenesis of IBD, is increased in inflamed LP of Crohn's disease.25 However, serum IFN-γ concentration in Crohn's disease was lower than that in the control in the present study (Fig. 4G). The low serum IFN-γ concentration in this study might explain one of the reasons anti-IFN-γ therapy failed to treat Crohnn's disease in Reinisch et al.'s trial.26 IL-21 is a pleiotrophic cytokine that has a central role in regulating effector functions of T cells, NK cells, and B cells.26 IL-21 is produced predominantly by CD4+ T cells and NKT cells.27 IL-21 can inhibit the differentiation of naïve CD4+ T cells to Treg and Th1.2728 IL-21 can also promote the differentiation of naïve CD4+ T cells to Th2, Th17, and Tfh cell.2728 IL-33 as one member of IL-1 family is a proinflammatory cytokine produced by intestinal epithelial cells and sub-epithelical myofibroblasts.11 It has been reported that the production of IL-33 of intestinal epithelial cells is increased in patients with ulcerative colitis, but not in patients with Crohn's disease.29 In our study, serum concentration of IL-33 in the normal control was higher than that in Crohn's disease or ulcerative colitis (Fig. 4F). The role of IL-33 has not been exactly evaluated yet in humans. The secretory function of IL-33 of intestinal epithelial cells might be decreased in the IBD patients. Further research could be performed.
Our results revealed that the serum concentration of IL-6 in Crohn's disease was significantly higher and tended to be higher in ulcerative colitis than that in the control (Fig. 4B). IL-6 is produced by macrophages, fibroblasts, and T cells.10 It is an important proinflammatory cytokine in IBD. It promotes differentiation of naïve CD4+ T cells to Th17 cell under the presence of TGF-β.1112
Serum concentrations of IL-17A and IL-22 in ulcerative colitis were significantly higher than those in Crohn's disease (Fig. 4C and E). IL-17A is a major pro-inflammatory cytokine and signature cytokine of Th17 cell.1112 We anticipated higher concentration of IL-17A in Crohn's disease and ulcerative colitis than that in the control. However, our results showed lower concentrations of IL-17A in Crohn's disease than that in the control and ulcerative colitis. This finding may suggest that Crohn's disease and ulcerative colitis have different pathogenesis mechanisms associated with Th17 cells. Strober et al.12 have reported the major role of Th1 cell and Sakuraba et al.30 reported the minor role of Th17 cell in the pathogenesis of Crohn's disease. Hueber et al.31 have demonstrated that Sekukinumab, a human anti-IL-17A monoclonal antibody, has no efficacy in Crohn's disease.
TNF-α has been identified as a key cytokine in IBD pathogenesis. Contrary to authors' anticipations, we could not find the difference of serum TNF-α concentration between the control and IBD groups (control group 21.5 ± 11.1 pg/mL; Crohn's disease group 21.6 ± 13.7 pg/mL; ulcerative colitis group 16.3 ± 6.8 pg/mL; P = 0.917). This finding might be caused by the fact that TNF-α is a cytokine secreted by innate immune cells located at tissue levels and most of that could be distributed to the inflamed tissues.
The limitations of our study are as follows. First, numbers of enrolled patients and normal control were small. The mean age was different between the control and IBD groups. Although the mean age of the control group was younger than that of the IBD group, laboratory conditions might not be different between the two groups because all subjects were in puberty. Unfortunately, cytokine analysis was only done for serum, not for both serum and intestinal mucosa.
In conclusion, mucosal immunity analysis showed increased FOXP3+ Treg cells in the LP of subjects with Crohn's disease. The paradoxical increase of FOXP3+ Treg cells in inflamed mucosa of Crohn's disease may not play a crucial role in the pathogenesis of Crohn's disease. However, there is still a possibility that a paradoxical increase of FOXP3+ Treg cells might reflect dysfunction of the cells indirectly. Th17 cell polarizing and signature cytokines were decreased in the serum of subjects with Crohn's disease, but increased in those with ulcerative colitis. This finding may suggest that Th17 cells do not have a crucial role in the pathogenesis of Crohn's disease. The complex crosstalk among several types of CD4+ effector T cells might play major roles in the chronicity of inflammation of IBD.
 XML Download
XML Download