

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
IgE-mediated anaphylaxis has been widely studied and is thought to be the main anaphylactic pathway. However, increasing evidence from animal studies supports the presence of a second pathway [1]. Such IgE-independent mechanisms can be mediated by IgG antibodies or complement proteins [23]. In the IgG-dependent pathway, macrophages and IgG, rather than mast cells and IgE, are the main immune components involved, and platelet-activating factor (PAF), rather than histamine, is the main mediator released [4].
In mice, the IgG-dependent pathway requires proportionately higher amounts of antibody and antigen than the IgE-mediated pathway [15]. High doses of allergen can induce IgG-mediated anaphylaxis by forming complexes that activate macrophages and basophils through the Fcγ receptor (FcγR) III. In a mouse model, this mechanism acted in concert with neutrophil activation resulting from the interaction between allergen-specific IgG2 and FcγRIV on cells. PAF was also the predominant mediator [6].
In humans, IgG receptors can activate PAF secretion by macrophages and neutrophils [17] and PAF can activate mast cells in vitro [8]; thus, PAF may contribute to human anaphylaxis [9]. IgG-mediated immune reactions in humans were observed following administration of certain drugs, including monoclonal antibodies (such as omalizumab and infliximab), von Willebrand's factor, and dextran. Given the increased use of different monoclonal antibodies in the clinical practice for treating immune disorders, an increase in this type of IgG-mediated anaphylaxis may occur [4].
The IgE-independent mechanism is clinically indistinguishable from IgE-mediated anaphylaxis [10]. To detect suspected drug-induced anaphylaxis, in vivo drug provocation testing is not recommended due to the risk of inducing a harmful reaction. Instead, whether such infusion reactions represent drug-induced anaphylaxis should be determined in vitro [11]. However, current in vitro tests for confirming the culprit drug mainly target specific IgE (sIgE) or basophils, through sIgE quantification, measurement of released mediators, such as tryptase and histamine, by mast cells, and basophil activation testing (BAT) [10].
Therefore, new tests for detection of the IgE-independent mechanism, particularly whether and when IgG-mediated anaphylaxis occurs, are urgently needed. We investigated the action of circulating neutrophils in peripheral blood. Neutrophils destroy microorganisms by producing reactive oxygen species as part of normal host defenses against infectious diseases [12].
The aim of this study was to develop an assay for diagnosis of IgG-mediated anaphylaxis. We investigated whether allergens trigger neutrophil oxidative burst (OB) and whether neutrophil OB occurs due to allergen-specific IgG (sIgG). Crude extracts of Dermatophagoides farinae (Der f) (American house dust mite) were employed as the allergen and neutrophil OB was measured by a dihydrorhodamine 123 (DHR 123) assay using flow cytometry.
Go to : 
MATERIALS AND METHODS
Subjects
A total of 31 subjects were selected according to the following criteria: 1) subjects who visited the allergy clinic of the Kyungpook National University Hospital (Daegu, Korea) and 2) subjects for whom multiallergen testing was conducted by the Department of Laboratory Medicine. The male to female ratio of the enrolled subjects was 16/15 and the median age was 44 years (3–76 yr). Lithium heparin tubes were used to collect whole venous blood samples for multiallergen testing. Samples that remained from these tests were used in the study. All participants provided informed written consent and the study was approved by the institutional review board. Experiments were performed within 4 h after blood collection.
Flow cytometry
A FACSCalibur flow cytometer equipped with the blue laser (488 nm) was calibrated daily using CaliBRITE beads and FACSComp software (BD Biosciences, San Jose, CA, USA). At acquisition, using CellQuest Pro software (BD Biosciences, San Jose, CA, USA), forward and side light scatter (FSC and SSC, respectively) were linearly amplified, whereas all fluorescence was logarithmically amplified.
DHR assay by flow cytometry
Leukocyte suspensions were separated from whole blood using an overlay procedure [13]. Briefly, 500 µL of whole blood was carefully overlaid onto 1.0 mL of Histopaque-1077 (Sigma-Aldrich, St. Louis, MO, USA) in a 1.5 mL microcentrifuge tube. The tube was left standing at room temperature for 20 min for red blood cell sedimentation. The supernatant (250 µL of plasma containing leukocytes) was carefully aspirated with a micropipette.
The isolated leukocyte suspension of each subject was divided into two fractions (“stimulated” and “unstimulated”), with a volume of 60 µL per tube, to which 2.0 µL of thawed and diluted DHR 123 (Sigma-Aldrich; final concentration = 375 ng/mL) were added [1214]. Both tubes were incubated at 37℃ for 5 min in shaking water bath to allow loading of DHR 123 into the cells, and the fraction to be stimulated (or the “stimulated” fraction) was spiked with a patch (diameter of −1 mm) of Der f extract (Institute of Allergy of Yonsei University College of Medicine, Seoul, Korea). Both tubes were subsequently incubated for 15 min at 37℃ in shaking water bath. Next, 125 µL of ethylenediaminetetraacetic acid (EDTA)-phosphate buffered saline (PBS) (0.01 M EDTA in PBS) at 4℃ were added to the tubes to stop the reaction and the samples were immediately submitted to flow cytometry analysis. The fluorescence of rhodamine 123 was measured using an FL1 (fluorescence 1) detector with a 530/30 bandpass filter.
The mean fluorescence intensity (MFI) of rhodamine 123 and the mean FSC of neutrophils were determined (Fig. 1). Two regional gates were used for neutrophils: one in the FSC/SSC plot and the other in the rhodamine 123/SSC plot. The MFI value and mean FSC value were expressed as the geometric means of the main portion (M1 or M2) of the peak in the rhodamine 123 histogram and in the FSC histogram of gated neutrophils, respectively. The change in MFI for unstimulated and stimulated neutrophils was expressed as the neutrophil oxidative index (NOI), which was calculated using Equation 1:
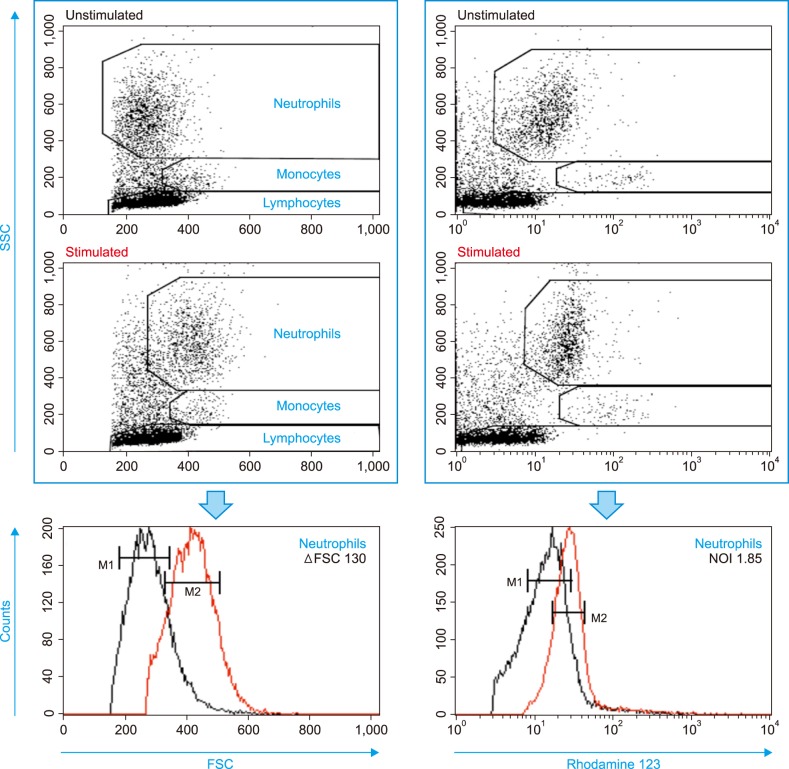 | Fig. 1Flow cytometry data acquisition and analysis for oxidative burst in response to Der f extract. Calculations of NOI and ΔFSC are described in the Materials and Methods section.Abbreviations: Der f, Dermatophagoides farinae; FSC, forward scatter; MFI, mean fluorescence intensity; NOI, neutrophil oxidative index; SSC, side scatter.
|
The change in FSC (ΔFSC) was calculated using Equation 2:
The cutoff value for distinguishing positive from negative results was 1.20 for NOI and 50 for ΔFSC.
CD63 assay
A BAT was performed by three-color flow cytometry [15]. For basophil priming, interleukin-3 (Sigma-Aldrich) was added to 100 µL of whole blood and incubated for 10 min at 37℃. This primed whole blood was spiked with a patch of Der f extract, and the reaction mixture was incubated for 30 min at 37° in shaking water bath. Samples were marked with the fluorescently labeled antibodies CD123 phycoerythrin-cyanine 5, CD203c allophyocyanin and CD36 fluorescein isothiocyanate for 30 min at 4℃. Following red blood cell lysis with 3.0 mL lysing solution and a single washing step, the remaining pellet was resuspended in PBS for flow cytometry data acquisition (all from BD Biosciences). For the CD63+ proportion of basophils, the cutoff value for distinguishing positive from negative results was 3.0%.
Determination of Der f-sIgE, Der f-sIgG (total), and Der f-sIgG4 levels
The level of Der f-specific (Der f-s) IgG (i.e., Der f-sIgG) antibodies (total) in the plasma samples was measured by enzyme-linked immunosorbent assay using RIDASCREEN Specific IgG kits and discs of Der f (R-Biopharm AG, Darfmstadt, Germany), with a limit of detection (LOD) of 2.5 µg/mL, and the levels of Der f-sIgE or Der f-sIgG4 antibodies were measured by a chemiluminescent immunoassay using ImmunoCAP Specific IgG (Thermo Fisher Scientific, Waltham, MA, USA), with an LOD of 0.1 µg/mL.
Statistical analyses
Statistical analyses were performed using SPSS Version 19.0 (SPSS, Inc., Chicago, IL, USA). The two groups were compared using regression analysis or Wilcoxon signed rank test. P values <0.05 were considered statistically significant. The results were expressed as median values (range).
Go to :
RESULTS
Time course of ΔFSC and NOI following activation with Der f extract
The leukocyte suspension was isolated using an overlay method for minimal manipulation. In unstimulated samples (not spiked with Der f extract), a time-dependent increase in the rhodamine 123 fluorescence of neutrophils was observed (Fig. 2A).
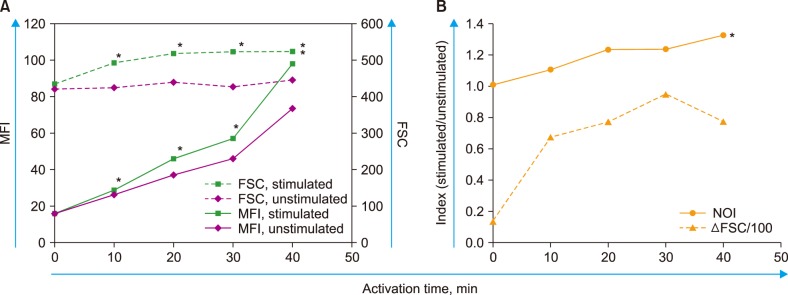 | Fig. 2Time course experiments showing effects of activation period on neutrophil reaction against Der f extract. Measurements were performed from 0 min (just prior to addition) to 40 min after adding Der f extract in 10 min intervals (total of five times). The typical results of four separate experiments are shown. (A) Raw data of FSC and MFI values. The difference between stimulated values and unstimulated values was significant (P<0.05) starting at 10 min activation for both FSC and MFI. Asterisks indicate significant stimulated values. (B) Calculated indices (ΔFSC and NOI). The linearity of Der f-sNOI values according to the activation time was significant (P<0.01, asterisk), which was not observed for the ΔFSC values.Abbreviations: Der f, Dermatophagoides farinae; FSC, forward scatter; MFI, mean fluorescence intensity; NOI, neutrophil oxidative index; SSC, side scatter.
|
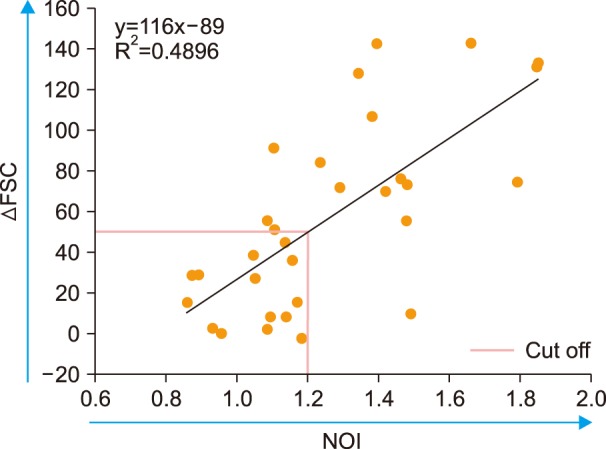
The stimulated samples (spiked with Der f extract) showed a faster increase in MFI than unstimulated samples, with a time-dependent increase observed in Der f-sNOI (Fig. 2B). ΔFSC reached a peak at 30 min of activation. When the linearity of these indices, according to the activation time from 0 min (prior to spiking) to 40 min, was evaluated, the linearity of the Der f-sNOI values was significant (P<0.01). However, that of ΔFSC values was not significant (P=0.118). Based on these results, 15 min was adopted as the activation period throughout this study. Measurements of the total 31 enrolled subjects revealed that ΔFSC was significantly correlated with Der f-sNOI (P<0.00005, Fig. 3).
A single subject (73-year-old woman) among 31 subjects enrolled showed a Der f-sIgG level below the LOD of 2.5 µg/mL (undetectable). This may be a case of life-long “non-responder,” i.e. who does not produce Der f-sIgG (see the DISCUSSION section). When excluding this potential non-responder against Der f, the level of Der f-sIgG was found to increase with age (P<0.05, Fig. 4A). However, no other significant relationship between Der f-sIgG and either Der f-sIgE (Fig. 4B) or Der f-sIgG4 (Fig. 4C) was observed.
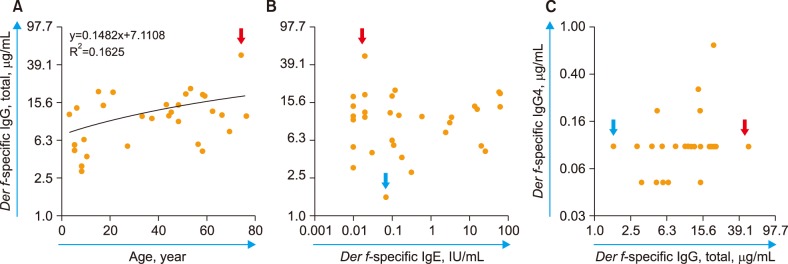 | Fig. 4Blood levels of Der f-sIgG (total) in 31 subjects and their correlation with (A) subject age (P<0.05); (B) Der f-sIgE level; and (C) Der f-sIgG4 level. In the panel C, IgG4 levels of five subjects with less than the limit of detection (0.1 µg/mL) were assumed to be 0.05 µg/mL for convenience. The red and blue arrows indicate extreme cases of Der f-sIgG levels (highest and lowest, respectively). In panel A, the case with the lowest (undetectable) Der f-sIgG levels (73-year-old woman, potential non-responder against Der f) was excluded from the regression analysis.
|
Der f-sNOI and Der f-sΔFSC values according to Der f-sIgG level
Neutrophil OB was nearly absent (NOI= −1) in both cases of the lowest and highest Der f-sIgG levels (<2.5 and 48.8 µg/mL, respectively). This resulted in a funnel-shaped distribution of the 31 subjects' dots in the Der f-sIgG vs. Der f-sNOI plot (Fig. 5A). When the lowest level case (non-responder against Der f) was excluded, the remaining subjects with detectable Der f-sIgG levels showed significant relationship between the two parameters (Fig. 5B); as the Der f-sIgG level increased, Der f-sNOI decreased (P<0.05). Der f-specific ΔFSC (Der f-sΔFSC) appeared to be negatively but not significantly correlated with Der f-sIgG levels (Fig. 5C).
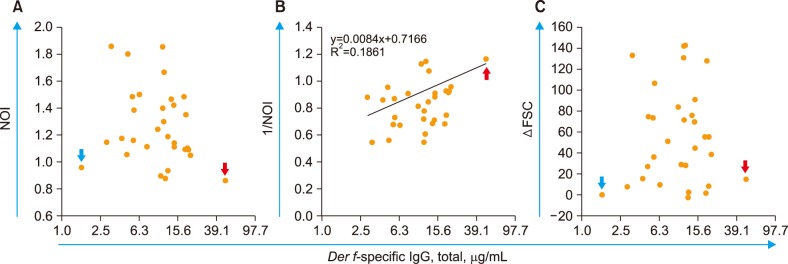 | Fig. 5Der f-sNOI and Der f-sΔFSC according to the Der f-sIgG level. The relationship between Der f-sIgG levels and (A) NOI; (B) 1/NOI (P<0.05); and (C) ΔFSC (P=0.201). The red and blue arrows indicate extreme cases of Der f-sIgG levels (highest and lowest, respectively). In panel B, the case with the lowest (undetectable) Der f-sIgG levels (73-year-old woman, potential non-responder against Der f) was excluded from the regression analysis.
|
Relationship between Der f-sNOI and other parameters
No significant relationship was found between Der f-sNOI and Der f-sIgG4, Der f-specific BAT (Der f-sBAT, percentage of CD63+ basophils), or Der f-sIgE (Fig. 6A–D). A decreasing trend in Der f-sNOI was observed with increasing age, but the results were not significant (P=0.157).
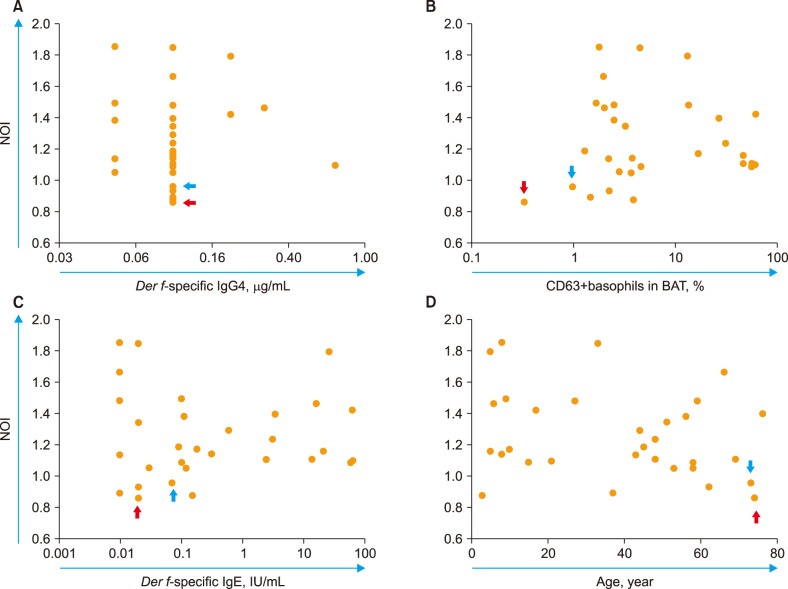 | Fig. 6Der f-sNOI and its relationship with (A) Der f-sIgG4; (B) Der f-sBAT; (C) Der f-sIgE; and (D) subject's age (P=0.157). No significant relationships were found. The red and blue arrows indicate extreme cases of Der f-sIgG levels (highest and lowest, respectively).Abbreviations: BAT, basophil activation test; Der f, Dermatophagoides farinae; NOI, neutrophil oxidative index.
|
Depriving leukocyte suspension of free preexisting Der f-sIgG
We selected four subjects who were previously positive for both Der f-sΔFSC and Der f-sNOI. Their leukocyte suspensions were washed three times with 3.0 mL of Roswell Park Memorial Institute medium to remove free preexisting Der f-sIgG. Neutrophil reactions were nearly absent in all the washed leukocyte suspensions, with a Der f-sΔFSC value (cutoff=50) of 2.0 (−6.0–15.0) and Der f-sNOI value (cutoff=1.20) of 1.05 (0.98–1.09). This suggests that neutrophil OB following the addition of Der f extract occurred because of free preexisting Der f-sIgG in the suspension.
Go to :
DISCUSSION
In this study, we verified that an allergen can trigger neutrophil OB via a preexisting allergen-sIgG immune complex, as measured by the DHR assay using leukocyte suspensions spiked with the allergen. This protocol is relatively simple and cost-effective.
In our protocol without red cell lysis, the leukocyte suspension was separated from red blood cells using an overlay method [13], which is suitable for rapid processing with minimal manipulation of leukocytes. We used Der f extract as an allergen to verify the role of allergen-sIgG in triggering neutrophil OB. In the Korean population, Der f is the most common sensitized allergen identified by the skin prick test (positivity of 40.9%) and ImmunoCAP (36.8%) [16].
Shortly after adding a tiny patch of Der f extract, a higher Der f-sIgG level was associated with a lower Der f-sNOI value. This result supports a previous study demonstrating that a high level of mite-sIgG is protective against mite anaphylaxis [17]. As soluble Der f allergen is slowly but continuously released into the suspension plasma, the blocking effect [5] of free Der f-sIgG is dominant during the initial activation period. However, this effect is shortly exhausted by released excessive allergen, which consumes free sIgG over an extended period. These immune complexes eventually attach to Fcγ receptors on neutrophils [181920] to trigger oxidative burst, which can be measured as Der f-sNOI (see Fig. 2: time course of neutrophil activation with Der f extract). In the leukocyte suspension separated by the overlay method, neutrophils were activated by themselves by slowly increasing their rhodamine 123 fluorescence, even when on stand-by status. Thus, the next steps of our protocol should be performed immediately after leukocyte separation.
In addition to the role of preexisting Der f-sIgG in neutrophil OB in our study, the direct effect of the Der f extract on neutrophils is another probable mechanism, similar to the effects observed for Hevein [21]. However, the latter mechanism is not likely in our study for two reasons: 1) a subject with undetectable Der f-sIgG (<2.5 µg/mL), who was a potential “non-responder” [22] to Der f, showed a Der f-ΔFSC of −0 and Der f-NOI of −1 (almost no neutrophil reaction); and 2) washed leukocyte suspensions from which Der f-sIgG was removed showed the same results.
We investigated the relationship between Der f-sNOI and parameters other than Der f-sIgG, Der f-sIgG4, Der f-sBAT, Der f-sIgE, and subjects' age. No parameters were significantly correlated with Der f-sNOI. The insignificant but decreasing trend in Der f-sNOI values with age may be related to the increased Der f-sIgG levels during aging.
Upon in vivo basophil activation, granules fuse with the plasma membrane and their contents are released in the extracellular environment within minutes, which can induce a chain reaction of neutrophil OB [232425]. We investigated whether this chain reaction occurs in our in vitro model using a leukocyte suspension and we did not find a significant correlation between Der f-sBAT and Der f-sNOI in the 31 subjects assessed (Fig. 6B). This finding suggests that neither in vitro basophil activation nor subsequent neutrophil OB occurs when our protocol is used.
Our protocol may be applicable for in vitro diagnosis of IgG-mediated anaphylaxis due to intravenously infused drugs or for detection of anti-drug antibodies that neutralize the biological activity of therapeutic proteins [26]. For this purpose, the amount of drug added should be sufficient to overcome the initial blocking effect of free sIgG. In contrast, when the allergen amount is optimized to reflect the blocking effect of free sIgG, this assay may be useful to assess the immunotherapy effectiveness.
We conclude that allergens can trigger neutrophil OB via preexisting allergen-sIgG and that neutrophil OB can be easily measured in a DHR assay using a leukocyte suspension spiked with the allergen. These findings can be used for in vitro diagnosis of IgG-mediated anaphylaxis induced by intravenously infused drugs.
Go to :
 XML Download
XML Download