

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Streptococcus pneumoniae is a gram-positive diplococcus that causes pneumonia and otitis media as well as severe invasive pneumococcal diseases (IPDs), such as sepsis and meningitis.1 The impact of pneumococcal infection is greatest among young children, the elderly, and subjects with predisposing conditions, such as immunosuppression.2 The polysaccharide (PS) capsule is the primary virulence factor for pneumococci; it shields the bacteria from host phagocytes and plays a significant role in immunogenicity.3
The development of an effective pneumococcal vaccine is crucial for reducing the incidence of infection worldwide.456 Two types of pneumococcal vaccines are currently available: pneumococcal polysaccharide vaccines (PPVs) and pneumococcal conjugate vaccines (PCVs).78 A 23-valent PPV (PPV23) that targets serotypes 1, 2, 3, 4, 5, 6B, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 17F, 18C, 19A, 19F, 20, 22F, 23F, and 33F was approved to prevent pneumococcal infections in subjects over 2 years of age with predisposing factors. Since PPV23 is not effective for young children (< 2 years of age), a 7-valent PCV (PCV7; serotypes 4, 6B, 9V, 14, 18C, 19F, and 23F) was developed910 and has significantly reduced the incidence of IPDs.1112 A 10-valent PCV (PCV10; PCV7 serotypes plus 1, 5, and 7F) and 13-valent PCV (PCV13; PCV10 serotypes plus 3, 6A, and 19A) were introduced in 2010.1314
The epidemiology of pneumococcal serotypes constantly changes owing to natural fluctuations15 and selective pressure from antibiotics and vaccines.161718 Routine vaccination has been influential in the reduction of IPDs, particularly those attributed to vaccine-related serotypes.19 However, reduction in vaccine-type IPDs has often been accompanied by an increase in IPDs from non-vaccine serotypes.20 PCV10 and PCV13 were necessary to prevent infection by serotypes that were not included in PCV7. Since PCV13 was introduced worldwide, various examples of serotype replacement have been reported in England, Wales, Japan, Israel, North America, and France.202122232425 This emergence of additional pneumococcal serotypes has led to the development of vaccines with broader strain coverage, such as 15-valent PCV (PCV15; PCV13 serotypes plus 22F and 33F).26
Vaccine-induced protective immunity is measured based on serum IgG antibody titrations by enzyme-linked immunosorbent assays (ELISAs)27; however, elderly patients with high pneumococcal antibody levels continue to show susceptibility to pneumococcal infection, indicating the presence of nonfunctional antibodies.32829 Therefore, an assay to evaluate the protective capacity of pneumococcal anticapsular antibodies is needed.28
Traditional antibodies directed to pneumococcal capsular PS antigens protect the host by opsonization, which marks the bacterium for destruction by phagocytes. As such, opsonophagocytic assays (OPAs) are widely accepted as a reference method to determine the protective capacity of pneumococcal antibodies.28 Since pneumococcal vaccines contain multiple serotypes, a comprehensive evaluation of vaccine immunogenicity requires several serotype-specific OPAs, necessitating a large amount of serum. To overcome this limitation, a 4-fold multiplexed OPA (MOPA) using antibiotic-resistant pneumococci as target bacteria has become a practical high-throughput tool to test 13 serotypes.30 However, PPV23 and the more recent conjugate vaccine formulations contain additional serotypes outside of those analyzed by current MOPA assays. To address this issue, a MOPA for 26 pneumococcal serotypes was developed by Nahm and Burton in 2012. Here, we established and validated this expanded pneumococcal MOPA platform to measure antibodies against 11 additional serotypes included in PPV23, with the goal of performing this assay in our laboratory as a standard assay for vaccine evaluation.
METHODS
Bacterial strains
The 11 pneumococcal strains used in this assay are listed in Table 1 (provided by Moon H. Nahm, University of Alabama at Birmingham, Birmingham, AL, USA). Strains were selected for resistance to optochin (OREP), spectinomycin (SPEC), streptomycin (STREP), or trimethoprim (TREP) (Table 1). To create working stocks, ~10 µL of each bacterial master stock was streaked onto blood agar plates (Hanil Komed, Seongnam, Korea) and cultured overnight at 37°C with 5% CO2. Bacterial colonies were subcultured in filter-sterilized Todd-Hewitt broth with 0.5% yeast extract (THY; Becton Dickinson, Sparks, MD, USA) in a 37°C water bath until reaching log-phase growth (OD600 = ~0.8). Multiple working assay stocks were then cryopreserved with ~15% glycerol and stored at −80ºC until use.

Table 1
Bacterial strain composition and antibiotic resistance of opsonization assay groups
| Assay groups | SPEC | OREP | STREP | TREP |
|---|---|---|---|---|
| A | SPEC6Ca | OREP3a | STREP33F | TREP22F |
| B | SPEC6Da | OREP10A | STREP8 | TREP12F |
| C | SPEC9N | OREP17F | STREP2 | TREP11A |
| D | SPEC20B | OREP17Fa | STREP2a | TREP15B |
SPEC = spectinomycin-resistant, OPEP = optochin-resistant, STREP = streptomycin-resistant, TREP = trimethoprim-resistant.
aStrains included to maintain an effector-to-target ratio of 200:1.
![]()
Assay stocks were deemed acceptable when the following criteria were met: 1) purity, as determined by streaking on blood agar plates and overnight incubation; 2) resistance to the appropriate antibiotic at a 2× selection concentration and sensitive to the other three antibiotics at 0.5×; and 3) an assay dilution factor of ≥ 103 to yield the ideal number of colonies per spot (80–150 CFU/spot).
Sera and antibodies
Three quality control (QC) serum pools were prepared by mixing equal volumes of serum from four healthy young adults (12 total donors, aged 22–34 years) 1 month after vaccination with PPV23 (Pneumovax; Merck & Co., Inc., Whitehouse Station, NJ, USA). The serum pools were expected to have variable opsonic indices (OIs) for each serotype.
A total of 42 serum samples were collected from healthy adults aged 20–50 years. Of these samples, 35 samples were obtained 14 to 27 days after immunization with PPV23 (Prodiax23®; Merck & Co., Inc.) by intramuscular injection (group 1). Seven samples were obtained from subjects immunized previously (46 to 123 months prior) with PPV23 (Prodiax23®) (group 2). OIs for 11 additional non-PCV13 serotypes were studied for 42 serum samples. The accuracy of MOPA was studied using 20 selected sera because of the limited volume for analysis.
Collected samples were stored frozen at −70°C until testing. All serum samples were heat-inactivated at 56°C for 30 minutes to inactivate endogenous complement activity and confirmed as antibiotic-free in test cultures with a rough pneumococcal strain (R36A).
Rabbit complement and fetal bovine serum (FBS)
One technical difficulty with MOPAs is the nonspecific bacterial killing by the rabbit serum used as a source of complement. As such, multiple lots of 3–4-week-old baby rabbit complement (PelFreeze Biologicals; Rogers, AR, USA) were screened for potency and nonspecific killing defined as: 100 × (1 − [CFU from rabbit serum/HL-60 reaction]/[CFU from heat-inactivated rabbit serum/HL-60 reaction]). FBS (HyClone, Logan, UT, USA) was heat-inactivated and screened in OPA reactions. Lots with equal to or less than 30% nonspecific killing for all 11 serotypes in the absence of human serum were deemed acceptable for use.
HL-60 cells
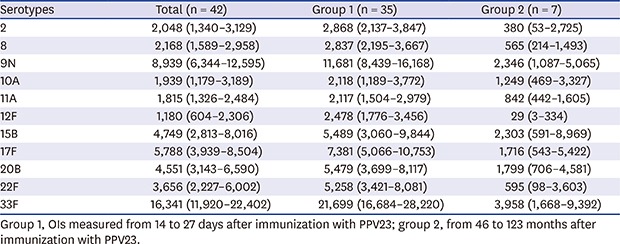
An HL-60 promyelocytic leukemia cell line derived from a 36-year-old female patient was obtained from American Type Culture Collection (CCL-240; American Type Culture Collection [ATCC], Manassas, VA, USA).31 Cells were expanded and cryopreserved immediately upon receipt. Cell aliquots were thawed and propagated in RPMI 1640 media supplemented with 10% FBS and 1% l-glutamine. To generate phagocytes for the OPA, HL-60 cells were differentiated for 5 days in the presence of 0.8% dimethylformamide at a starting density of 4 × 105 cells/mL. Differentiated HL-60 cells have been shown to express the markers of mature neutrophils when differentiated into granulocytes.3233 It was important to demonstrate the presence of these receptors under the differentiation conditions used in this particular assay, since complement-dependent opsonophagocytosis depends on the presence of the appropriate cell receptors on the effector cell. Differentiated HL-60 cells were considered acceptable as effector cells if the following criteria were met: 1) viability ≥ 90% (trypan blue), 2) CD35, also known as CR1 or C3b receptor which presents on granulocytes, expressed on ≥ 55% of cells, 3) CD71, presenting on proliferating cells as transferring receptor, expressed on ≤ 20% of cells, 4) ≤ 25% apoptotic cells, defined as Annexin V+/PI– by flow cytometry using a FACS Calibur (Becton-Dickinson, San Jose, CA, USA).
MOPA
Opsonic activity was evaluated by MOPA as previously described.30 Briefly, frozen aliquots of target pneumococci were thawed, washed twice, and diluted to the proper bacterial density (~5 × 104 CFU/mL for each serotype). Serum samples were plated in 3-fold serial dilutions (20 µL/well) in round-bottom 96-well plates (Corning Inc., Corning, NY, USA). Then, 10 µL of bacterial suspension was added to each well in duplicate and incubated for 30 minutes at room temperature (25°C) with shaking (Mini Orbital Shaker; Bellco Biotechnology, Vineland, NJ, USA) at 700 rpm. Next, 10 µL of rabbit complement and 40 µL of differentiated HL-60 cells (4 × 105 cells) were added to each well. Plates were incubated in a tissue culture incubator (37°C, 5% CO2) with mechanical agitation at 700 rpm for 45 minutes and transferred to ice for 10–15 minutes. Next, an aliquot of the final reaction mixture (10 µL) was spotted onto four different THY agar plates (THY in 1.5% agar). When the fluid was absorbed into the agar, an equal volume of overlay agar (THY with 0.75% agar and 25 mg/L 2,3,5-triphenyltetrazolium chloride) containing one of the four antibiotics was added to each plate. After overnight incubation at 37°C, bacterial colonies were counted using the National Institute of Standards and Technology (NIST) Integrated Colony Enumerator (NICE), freely available in the US NIST (Gaithersburg, MD, USA; http://www.vaccine.uab.edu/NICE.pdf), and analyzed using Opsotiter 3.0 (http://www.vaccine.uab.edu). The OI was defined as the serum dilution that killed 50% of bacteria by linear interpolation, as previously described.30
Assay validation
MOPAs were subjected to accuracy, specificity, and reliability analyses to validate their application as an analytic methodology to evaluate pneumococcal vaccine immunogenicity. Since there is no standard or reference serum with assigned OPA values, OIs from the MOPA were compared with those from single-serotype OPAs (SOPAs) with 20 serum samples from vaccinated adults to determine assay accuracy. Assay specificity was examined by neutralizing QC sera in reactions with homologous PS (ATCC), three heterologous capsular PSs, or with no PS before being evaluated separately in the OPA. In an additional assay for serotype 20B, another capsular PS of serotype 20, known as serotype 20B, was used (SSISP20/2; Staten Serum Institute [SSI], Copenhagen, Denmark).34 Equal volumes of diluted serum and PS were mixed and incubated at room temperature (25°C) for 15 minutes. The sera were then serially diluted with opsonization buffer B (OBB), and the reactions were performed as described above. Intra-assay variation for the 11 serotypes was determined based on the coefficient of variation (CV) for a serum pool estimated 10 times in a single assay run. Similarly, inter-assay precision was estimated by testing several sera in 10 independent experiments over a 2-week interval. For these studies, the same lots of target bacteria and complement were used for each assay.
OPA application to sera samples
To apply this expanded MOPA to immune sera, the functional antibodies against 11 serotypes of pneumococcus (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20B, 22F, and 33F) were evaluated in 42 sera immunized with PPV23 previously. Sera samples were assigned to two groups according to their intervals between immunization and blood sampling such as group 1 and group 2.
Statistical analysis
Correlation analysis of OIs from MOPAs and SOPAs was performed to assess accuracy, and correlation coefficient (r) values for each pneumococcal serotype were calculated by the Pearson's correlation coefficient using SPSS statistical software (version 23.0; IBM Corp., New York, NY, USA). The overall mean ± standard deviation (SD) and the CV of OIs were calculated to assess precision. OIs of 42 samples were reported as geometric mean indices (GMIs) and 95% confidence interval. GMIs between two groups were compared using the Mann-Whitney U nonparametric test on the 5% significance level.
Ethics statement
The study protocol was reviewed and approved by the Institutional Review Board of Ewha Womans University Mokdong Hospital (ECT 11-13-43 and EUMC 2016-01-012-001). The study was conducted in accordance with clinical best-practices (national regulations and ICH E6) and the principles of the Helsinki Declaration. Written informed consent was obtained from all participants following a detailed explanation of the study.
RESULTS
Validation of bacterial stocks and HL-60 cells
Antibiotic-resistant strains were assessed with respect to resistance and sensitivity to the four antibiotics utilized in the MOPA. As expected, all 11 resistant strains showed 20% or less diminished growth when cultured in the presence of a 2× concentration of the appropriate antibiotic, but were sensitive to the other three antibiotics at half the concentration used in the assay with reduction rates ranging from 96% to 100% (data not shown). All 11 bacterial strains met the assay dilution cut-off of ≥ 103.
The phenotype of HL-60 cells that differentiated into phagocytes over 5 days was assessed by flow cytometry (Fig. 1). The percentage of apoptotic cells (Annexin V+/PI–) was 20.2%, which met the ≤ 25% acceptance criteria. Moreover, 83.0% and 11.8% of cells expressed CD35 (acceptance criteria, ≥ 55%) and CD71 (acceptance criteria, ≤ 20%), respectively, indicating sufficient phagocytic differentiation.
Nonspecific killing
The nonspecific killing attributed to complement and FBS ranged from −14% to 12% and from −16% to 9%, respectively, which met the necessary criterion of equal to or less than 30% (Supplementary Table 1).
Assay validation
Accuracy
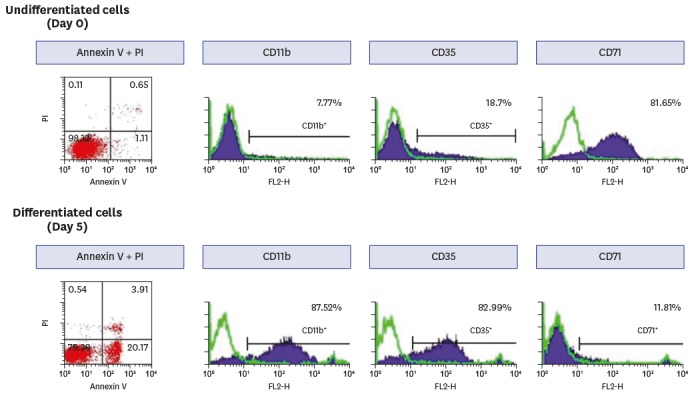
MOPA accuracy was assessed by correlation analyses with data generated in SOPA reactions. The data for all 11 serotypes are shown in Fig. 2, along with the identity line (dotted line) and lines showing two-fold deviation (solid lines) from identity for each serotype. The results of all 220 data points (11 serotypes × 20 samples) obtained from each format agreed with only one exception, data point (for serotype 10A) that deviated more than two-fold (i.e., outside of the two solid lines).
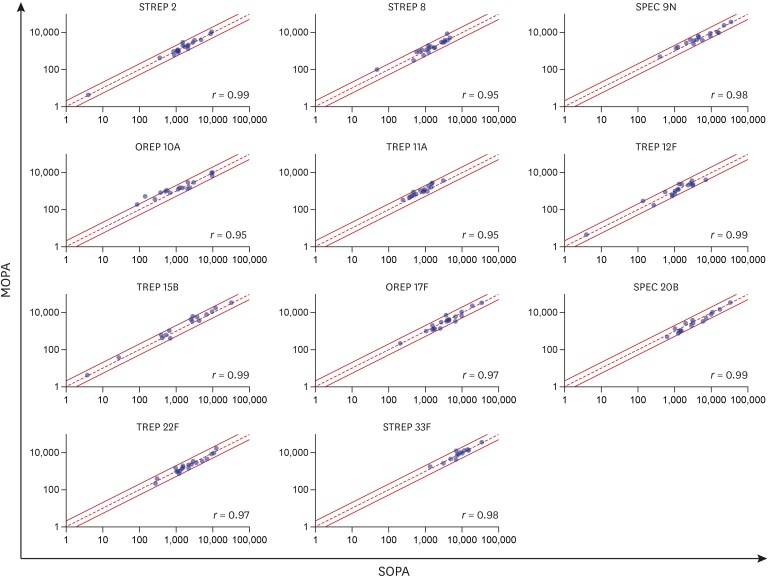
Fig. 2
Correlation analysis of MOPA and SOPA OIs. Scatter plots of OIs for all 11 serotypes with lines indicating the 45° reference (dotted) and two-fold deviations (solid). The correlation coefficient (r) value for the relationship between SOPA (x-axis) and MOPA (y-axis) is shown on each plot.
MOPA = multiplexed opsonophagocytic assay, SOPA = single-serotype opsonophagocytic assay, OI = opsonic index, OREP = optochin-resistant, SPEC = spectinomycin-resistant, STREP = streptomycin-resistant, TREP = trimethoprim-resistant.
![]()
Specificity
MOPA specificity was determined by pre-absorbing a QC pool with homologous and heterologous capsular PSs and then testing reactivity in individual assays. This analysis revealed that opsonization was inhibited for all strains in the presence of homologous capsular PS, with the exception of SPEC20B (Fig. 3). As shown in Fig. 3, pre-incubation of the serum pool with serotype 20 PS (ATCC209-X from the ATCC) decreased the SPEC20B OI by only ~40%–50% when compared with the control reaction. Since these results showed the low inhibition of SPEC20B killing with homologous PS, the serum pool was pre-absorbed with another capsular PS of serotype 20 (SSISP20/2 from SSI). Pre-incubation with SSISP20/2 completely abrogated SPEC20B killing. Little or no OI inhibition was observed when test sera were pre-incubated with any of the three heterologous capsular PSs in the assay group, including SPEC20B.
Fig. 3
Specificity of the MOPA for 11 additional serotypes. The number of surviving CFU (y-axis) at various dilutions of serum (x-axis) pre-absorbed with heterogeneous PSs, homogenous PSs, or buffer alone as a control. The solid and dashed horizontal lines represent 0% and 50% killing, respectively.
MOPA = multiplexed opsonophagocytic assay, CFU = colony forming unit, PS = polysaccharide, Pn = Streptococcus pneumoniae, ATCC = American Type Culture Collection, SSI = Staten Serum Institute.
aSPEC20B partially inhibited serotype 20 PSs (ATCC), whereas serotype 20 PSs (SSI) completely inhibited opsonization.
![]()
Precision
Intra-assay variation for the 11 serotypes was assessed by calculating the CV of the 10 values obtained in a single assay run (Table 2). The CV for each serotype in all three pools was 30% or less and thus was deemed acceptable based on the guidelines established by the University of Alabama Birmingham MOPA protocol (www.vaccine.uab.edu). Similarly, inter-assay precision was estimated by testing several sera in 10 independent experiments over a 2-week interval (Table 2). For each QC pool, CVs were higher than the 30% cut-off for acceptance; however, all others were below this threshold and deemed reliable.
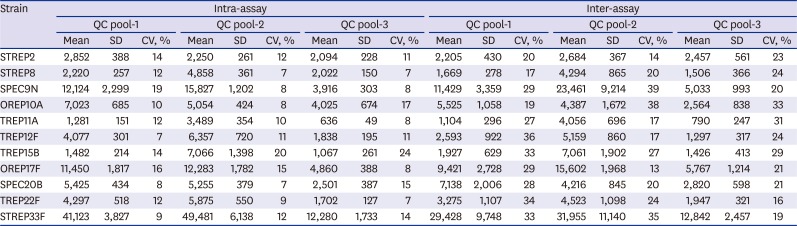
Table 2
Intra- and inter-assay precision of MOPA OIs for 11 bacterial serotypes
MOPA = multiplexed opsonophagocytic assay, OI = opsonic index, QC = quality control, SD = standard deviation, CV = coefficient of variation, STREP = streptomycin-resistant, SPEC = spectinomycin-resistant, OREP = optochin-resistant, TREP = trimethoprim-resistant.
![]()
OPA of immune sera
The median age of total volunteers was 35 years (range, 20–50 years), and that of group 1 and group 2 was 35 years (20–48 years) and 33 years (30–50 years), respectively. The mean ages were 35.0 ± 7.4 years for total, 34.5 ± 7.4 years for group 1, and 37.0 ± 8.5 years for group 2. The total number of males was 27, and 26 of them were assigned to group 1. The range between immunization and bleeding of group 1 was 14–22 days, and that of group 2 was 49–123 months.
The OIs obtained from forty-two clinical samples using this expanded MOPA are described in Table 3. The GMIs of group 1 for all serotypes were significantly higher than those of group 2 except for serotype 15B (P < 0.05).
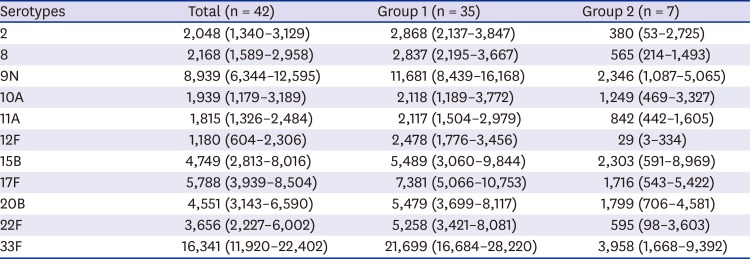
Table 3
Serotype-specific geometric mean OIs (95% CI) as measured by the MOPA for healthy adults (n = 42) immunized with the PPV23
Group 1, OIs measured from 14 to 27 days after immunization with PPV23; group 2, from 46 to 123 months after immunization with PPV23.
OI = opsonic index, CI = confidence interval, MOPA = multiplexed opsonophagocytic assay, PPV23 = 23-valent pneumococcal polysaccharide vaccine.
![]()
DISCUSSION
In this study, an expanded MOPA platform was established and validated to 11 additional target serotypes included in PPV23 (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, and 33F). Each assay component was prepared to satisfy the validation criteria. The bacterial stocks demonstrated resistance to specific antibiotics. However, they retained sensitivity to the other three antibiotics used in the assay. HL-60 cell differentiation into phagocytes was confirmed by phenotypic expression and viability analyses. Moreover, both complement and FBS were confirmed to have limited effects on nonspecific killing.
Since there is no OPA reference standard available, it was impossible to establish an independent marker of accuracy for MOPA.34 Rather, the results for each serotype were evaluated in a comparative analysis with the results obtained for SOPAs. The assays yielded similar data for all serotypes, with deviations of less than two-fold deviations for most samples. These findings confirmed that our MOPA had high accuracy.
An analysis of assay specificity showed that the MOPA was highly serotype-specific. i.e., the pre-absorption of test sera with homologous capsular PS resulted in significant decreases in OIs, while pre-absorption with heterologous capsular PSs resulted in negligible changes. One exception was serotype 20 (SPEC20B), which showed two distinct patterns of inhibition for ATCC serotype 20 PS (ATCC209-X) and SSI serotype 20 PS (SSISI20/2). This was consistent with a previous study that showed serologically different patterns for PS among serotype 20 strains.34 These were assigned to serotype 20A and 20B, and the biochemical, genetic, and serological characteristics of these subtypes were determined.35 Additionally, different antigenic properties were observed between SPEC20B and serotype 20 included in PPV23. An antigenic gap between vaccine strains and target strains of the MOPA should be considered when interpreting the immunogenicity results using the MOPA.
The low inhibition of STREP33F may be due to the relatively high OIs, rather than insufficient assay specificity. Thus, the 11 different serotype-specific PSs inhibited homologous bacteria, but not heterologous bacteria, thereby demonstrating high serotype specificity.
Under laboratory conditions, the expanded MOPA had intermediate precision, with CVs that were generally less than 30% for most of the 11 additional serotypes, suggesting adequate reproducibility in both intra- and inter-assay runs. However, high CVs (> 30%) were associated with specific test samples and serotypes (Table 2). Immunoassays based on biochemical antigen-antibody interactions usually show inter-assay CVs of less than 20%36; however, those based on biological functions are more complex and can exhibit high CVs.30343738 Functional assays are not based on simple antigen-antibody reactions and often utilize less-controllable biological agents, such as bacteria, cells, and complement. In a previous MOPA validation study, cross-reactive antibodies had higher CVs (> 50%) than those of vaccine-induced antibodies.34
ELISAs have been applied to evaluate pneumococcal vaccine immunogenicity for the 11 serotypes, since no standard references exist. However, studies have reported that ELISA is not recommended for evaluating immunogenicity in adults since adults are susceptible to pneumococcal infections despite having high quantitative levels of antibodies.29 Moreover, they suggested functional immune responses may yield better results than ELISA to discriminate low vaccine responders among the elderly.39 Notably, the assay established in the present study provides a means to assess pneumococcal antibody function in Korea. Lee et al.40 first demonstrated that PPSV23 induced functional immune responses to 11 non-PCV13 serotypes in Korean elderly by the expanded MOPA.
Recently, a new PCV15 was developed by Merck and includes the PCV13 serotypes as well as 22F and 33F, which account for ~10% of IPD cases in adults.41 Thus, PCV15 has significant medical and public health implications by providing broader coverage of the leading pathogenic serotypes.42 The results obtained from this study are expected to evaluate the functional antibody responses of PCV15 for serotypes 22F and 33F, and moreover for developing new pneumococcal PCVs.
To apply the established MOPA to the analysis of actual immune sera, we evaluated 42 immune sera. As expected, OIs of samples measured at 14 to 27 days after immunization with PPV23 (group 1) were significantly higher than those of samples measured at 46 to 123 months after immunization (group 2) against 10 additional target serotypes in PPV23. Though not showing significant differences, the OI to 15B serotype was higher in group 1.
This study had two limitations. First, MOPA accuracy was evaluated by comparing the results of both MOPAs and SOPAs owing to lack of standard or QC sera and their assigned value for each serotype. A recent study proposed that normalization of MOPA results from different laboratories using a reference serum (“007sp”) significantly reduced the interlaboratory variation.43 Thus, further studies using serum standards such as 007sp, would allow for a better measure of assay accuracy in future MOPA platforms. Moreover, inter-assay precision was examined over a short time period at a single center, and a multicenter analysis over an extended period using different batches of bacteria and complement may be useful. However, the results from this study supported the notion that the expanded MOPA is applicable to evaluate vaccine-induced antibodies. Consequently, the antibodies of additional 11 serotypes can be assessed for functional activity by the expanded MOPA. Since QC and reference sera for these serotypes are recently established,43 we believe that this assay could be applied to evaluate immune response of not only PPV23 but also newly developing PCVs, in the near future with more reliability.
In this study, we established a MOPA, which is available for 11 additional serotypes in PPV23. The expanded MOPA platform described herein provides a method to assess the full immunogenicity of PPV23 vaccination; this method was validated with respect to accuracy, specificity, and precision. Importantly, it may also be useful to evaluate future formulations of conjugate vaccines.
 XML Download
XML Download