

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Dapsone is a member of the sulfone family and has been used as an antibiotic [16]. To inhibit the growth of bacteria, dapsone disturbs the synthesis of dihydrofolic acid in bacteria by competing with para-aminobenzoic acid by acting on dihydropteroate synthetase [1617]. Dapsone has been used to treat multibacillary leprosy in combination with rifampicin and clofazimine [17].
Dapsone has been shown to have anti-inflammatory effects in addition to antibiotic activity, and two functional mechanisms have been suggested so far. The first is to modulate the production of inflammatory cytokines. Dapsone inhibits the secretion of interleukin-8 from lipopolysaccharide (LPS)-stimulated human bronchial epithelial cells [8] and the production of tumor necrosis factor alpha (TNF-α) from activated mononuclear cells [1]. The second mechanism suggested for the anti-inflammatory effect of dapsone is to inhibit myeloperoxidase (MPO) in neutrophils. MPO in neutrophils produces hypohalous acids involved in the microbicidal activity of neutrophils [3]. A previous study demonstrated that dapsone affected the function of neutrophils by blocking MPO in azurophilic granules; as a result, dapsone reduced the accumulation of hypohalous acid, a causative agent for tissue damage [9]. The effect of dapsone on neutrophil MPO is associated with its anti-inflammatory effects. Although these two mechanisms of dapsone have been studied, the anti-inflammatory effects of dapsone have not been fully elucidated yet.
In this study, we treated mouse bone marrow cells (BMs) with LPS to simulate the inflammatory reaction environment in osteomyelitis. LPS acts as prototypical endotoxin in tissues and binds to various receptors including cluster of differentiation 14 and Toll-like receptor 4 in many types of immune cells such as macrophages [12]. The binding of LPS triggers the production of pro-inflammatory cytokines, nitric oxide, and eicosanoids [14].
In the present study, we cultured mouse BMs in the presence of LPS and after treatment with dapsone. The levels of cellular activity, cytokine production, and reactive oxygen species (ROS) were analyzed in the study experiments.
Materials and Methods
Animals and reagents
C57BL/6 mice were purchased from OrientBio (Korea) and maintained in our animal facility. The present study used 8- to 12-week-old mice. All experiments using animal were performed according to the institutional guidelines of Jeju National University for laboratory animal use and care (IACUC No. 2015-0017). Dapsone and LPS purified from Escherichia coli O55 were purchased from Sigma (USA). For use, dapsone and LPS were dissolved in sterile phosphate buffered saline.
Preparation and culture of BMs
BMs were harvested from the femurs and tibias of C57BL/6 mice as described previously [10]. Any contaminated red blood cells in the samples were eliminated by applying ACK lysis buffer. BMs were filtered through a 70 µm cell strainer to obtain single cells. To culture BMs, RPMI1640 media containing 5% fetal bovine serum, 2 mM L-glutamine, and 100 U/mL penicillin/streptomycin (Thermo Fisher Scientific, USA) were used.
Measurement of cellular activity
The cellular activity of BMs was evaluated after treatment. Briefly, BMs were seeded at a concentration of 1 × 106 cells/mL in 96-well culture plates and treated with LPS and varying concentrations of dapsone. After culturing for 3 days, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT; Sigma) was added for 4 h (0.5 mg/mL) [7]. The viable cells generated insoluble violet crystals, and 100 µL/well of 10% sodium dodecyl sulfate solution was added for 2 h to dissolve the crystals. The optical density of the sample was measured at 570 nm by using a microplate reader (Multiskan FC; Thermo Fisher Scientific).
BM cell morphology
To observe alterations of cell morphology, BMs were seeded at a concentration of 1 × 106 cells/mL in 24-well culture plates and then treated with LPS and dapsone. After culturing for 3 days, the morphology of treated cells was photographed using an inverted microscope (IX70; Olympus, Japan) and a digital camera.
Measurement of cytokine amount
BMs (1 × 106 cells/mL) were established in 96-well culture plates and treated with LPS and dapsone at the indicated concentration. After 3 days, the supernatants were collected and used to determine the level of TNF-α, a representative pro-inflammatory cytokine. The cytokine concentration was measured by performing an enzyme-linked immunosorbent assay (ELISA) using a CytoSet antibody pair (Thermo Fisher Scientific) based on the manufacturer's manual.
Flow cytometry analysis
BMs were cultured for 3 days at a concentration of 1 × 106 cells/mL in 6-well culture plates and treated with 50 µg/mL dapsone and 1 µg/mL LPS. For flow cytometry analysis, the treated BMs were harvested and subjected to multiple assays. We measured cell size (forward scatter [FSC]) and cell granularity (side scatter [SSC]) of the treated BMs. To analyze cell death, both apoptosis and necrosis, the cells were stained with 0.25 µg/mL propidium iodide (PI; Sigma). To check the mitochondrial membrane potential (MMP) in the BMs, the cells were incubated with 10 µg/mL Rhodamine 123 (Sigma) for 30 min at room temperature. Additionally, to detect granulocytes, a specific antibody for granulocyte differentiation antigen 1 (Gr-1) was used as the primary antibody with streptavidin-fluorescein isothiocyanate as the secondary antibody (BD Biosciences, USA). To measure intracellular ROS level, the cells were treated with 10 µM dichlorofluorescein diacetate (DCFDA; Sigma). All stained cells were analyzed by using a FACSCalibur flow cytometer and CellQuest software (BD Biosciences).
Statistical analysis
The data from the MTT assay and ELISA are presented as mean ± SD values and were statistically analyzed by one-way analysis of variance followed by Tukey's multiple comparison test as provided by InStat (GraphPad Software, USA). A p value of < 0.05 was considered significant. The *, **, and *** symbols indicate p < 0.05, 0.01, and 0.001, respectively, compared to control values.
Results
Effects of dapsone on the cellular activity of LPS-treated BMs
To investigate the effects of dapsone on LPS-treated BMs, the cells were treated with LPS at 1 µg/mL and dapsone over a range of concentrations (0–100 µg/mL). Results of MTT assays demonstrated that LPS significantly enhanced the cellular activity of BMs compared to that in control BMs without LPS treatment (Fig. 1). Dapsone significantly decreased the cellular activity of LPS-treated BMs across a range of dapsone concentrations (12.5–100 µg/mL). These results indicate that dapsone can suppress the cellular activity of BMs that have been stimulated by inflammatory agents, such as LPS. In the following experiments (Figs. 2, 3, 4 and panel B in Fig. 5), the 50 µg/mL concentration of dapsone was used based on the drug's function and cytotoxicity.
Dapsone down-regulates activation of granulocytes by LPS
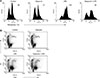
In the dot plots of panel A in Fig. 2, the region circled includes granulocytes, such as neutrophils, based on the relationship between FSC and SSC. LPS increased the percentage of the granulocytes, and dapsone markedly decreased the LPS increase in granulocytes. This result indicates that dapsone considerably decreases the number of activated granulocytes in an LPS-induced inflammation state. To further investigate if the region within the dot plots includes granulocytes, we employed Gr-1 antigen to detect granulocytes specifically. The BMs were stained with anti-Gr-1 antibody and plotted against SSC (panel B in Fig. 2). While the percentage of Gr-1 positive cells was 57% in control cells, LPS increased that level to 75%, while dapsone plus LPS decreased to 64%. The dot plots confirmed that LPS increases the percentage of granulocytes with enhanced granularity (SSC), whereas dapsone decreases the LPS-increased percentage of granulocytes. In panel B in Fig. 2, two regions can be detected based on the extent of the granularity (i.e., middle and high SSC, R1 and R2, respectively). Interestingly, dapsone decreased the percentage of granulocytes in R1 (middle SSC) in the LPS-treated BMs (panel B in Fig. 2).
Dapsone affects BM morphology
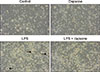
To observe the morphological changes in BMs produced by LPS and/or dapsone treatment, we obtained pictures by using an inverted microscope connected to a digital camera. There were no notable differences between the control and dapsone-treated BMs (Fig. 3). However, LPS treatment generated some markedly large BMs (Fig. 3; black arrows) after 3 days of treatment, and dapsone treatment decreased the size of the large BMs in LPS-treated BMs.
Dapsone diminishes MMP and increases cell death of LPS-activated BMs
To investigate how dapsone affects LPS-activated BMs, we measured MMP of the cells. For this purpose, a rhodamine 123 solution was used to stain BMs treated with LPS and/or dapsone. LPS enhanced the MMP of BMs, whereas dapsone profoundly decreased the LPS-increased MMP of BMs (panel A in Fig. 4). This indicates that dapsone can destabilize the structure of the mitochondrial double membrane. To investigate further the cellular status, PI solution was used to detect the percentage of dead cells. It is generally accepted that the cell size of dead or dying cells is gradually decreased. Thus, we defined the dot plot region with low FSC and high PI as one contains dead cells (panel B in Fig. 4). LPS decreased the percentage of dead cells from that in the control samples. However, dapsone increased the percentage of dead cells compared to those in both the control and LPS-activated BMs (panel B in Fig. 4). These results show that dapsone can induce cell death in BMs via disturbance of the MMP.
Effects of dapsone on production of TNF-α and ROS in LPS-activated BMs
Quantified ELISA specific for TNF-α was used to investigate whether dapsone may affect cytokine production of BMs treated with LPS. Despite the low concentration (25 µg/mL) treatment, dapsone significantly decreased the production of TNF-α in LPS-activated BMs by 54% (panel A in Fig. 5). This result indicates that dapsone can down-regulate the production of pro-inflammatory cytokines in BMs. We also investigated whether dapsone can modulate ROS in BMs; however, the results showed that dapsone did not change the level of ROS production in BMs (panel B in Fig. 5).
Discussion
Anti-inflammatory drugs include steroid and non-steroidal anti-inflammatory drugs that can disturb cyclooxygenase [513]. Dapsone is commonly used as an antibiotic to treat leprosy [1617]. Though its anti-microbial mechanisms have been studied, the role of dapsone in inflammation is not clear yet. To elucidate the mechanism of dapsone on inflammatory cells, we cultured BMs in the presence of LPS to simulate an inflammatory environment. LPS is a representative inflammatory agent and causing endotoxemia, a kind of sepsis in hosts [15].
The effect of dapsone on the cellular activity and viability of LPS-activated BMs was measured by using MTT assays and nuclear/cellular staining with fluorescent dyes. Dapsone consistently damaged the viability of LPS-activated BMs. Flow cytometry analysis revealed that dapsone controls LPS-generated inflammatory cells, especially granulocytes. While the percentage of Gr-1 positive cells was 57% in control, LPS increased that level to 75%, and LPS plus dapsone treatment decreased the LPS level to 64%. These results suggest that dapsone has immunomodulatory effects through down-regulation of the excessive ratio of inflammation-related cells in BMs produced by LPS.
The present study demonstrated that dapsone can decrease the percentage of granulocytes in LPS-activated BMs. This result may be related to agranulocytosis, one of the reported side effects of dapsone [26]. However, our study was performed in BMs with an excessive inflammatory response generated by LPS, not under normal conditions. Thus, the effect is different from agranulocytosis. Flow cytometry analysis demonstrated that LPS enhanced the MMP of BMs, whereas dapsone profoundly decreased the LPS-increased MMP of BMs. The observation that dapsone can inhibit the MMP in inflammatory cells or granulocytes has not been previously reported. The present results show that dapsone reduces both number and mitochondrial function of inflammatory cells when LPS is present.
To investigate further the anti-inflammatory effects of dapsone, we measured the production of TNF-α in LPS-activated BMs. TNF-α is a representative pro-inflammatory cytokine and is essentially produced during a variety of inflammatory reactions [411]. Interestingly, dapsone at a low concentration (25 µg/mL) significantly decreased the production of TNF-α by 54% from that in LPS-treated BMs. This result may broaden the uses of dapsone to include down-regulating the production of inflammatory cytokines.
Taken together, this study has demonstrated that dapsone can modulate the abundance of inflammatory cells, including granulocytes, in LPS-activated BMs by inducing cell death and down-regulating TNF-α production. Considering that dapsone is widely used as an antibiotic, and its safety is secured, our results indicate the possibility of developing it as an anti-inflammatory drug.



 XML Download
XML Download