

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Ischemia/reperfusion injury (IRI) and nephrotoxicants provoke inflammatory responses (1). Although inflammation is a protective mechanism to maintain homeostasis, it paradoxically contributes to tissue injury. Therefore, the resolution of inflammation is an essential step for organs, such as the kidneys, to repair themselves. While the mechanisms of inflammation following IRI are relatively well characterized, the mechanisms involved in the resolution of inflammation have not been thoroughly investigated.
Neutrophils, effector cells in innate immunity, rapidly infiltrate the kidneys at the early stage of reperfusion and blocking neurtrophil-endothelial interaction resulted in renoprotection in an animal model of IRI (2). Neutrophils are terminally differentiated cells with a short life span of less than several hours, rapidly infiltrate tissue, execute their innate immune function, die by apoptosis, and are cleared by macrophages. However, there have been no studies that convincingly demonstrate that apoptosis of neutrophils is critical in the resolution of inflammation or functional and histological recovery. In this study, we sought to determine the occurrence of neutrophil apoptosis and its role in the resolution of inflammation and in the recovery process by using Bax knockout (KO) bone marrow chimera in which only bone marrow derived cells are partially resistant to apoptosis.
In addition to neutrophil apoptosis, several recent studies demonstrated that tissue-infiltrating neutrophils can reenter the circulation by a process called reverse transendothelial migration (rTEM). Woodfin et al. (3) demonstrated the occurrence of rTEM of neutrophils after IRI, and also showed that junctional adhesion molecule-C (JAM-C) is an important negative regulator of rTEM. Therefore, in this study, we also evaluated whether rTEM also occurs in the recovery phase of IRI and whether rTEM contributes to the resolution of inflammation as well as to functional and histological recovery.
Collectively, in this study, we investigated the contribution of apoptosis, and rTEM to clearance mechanism of neutrophils and recovery following IRI.
MATERIALS AND METHODS
Animals
Six- to eight-week-old male C57BL/6 mice (20–25 g) were purchased from Orient (Seoul, Korea) and were allowed unrestricted access to water and chow before manipulation. Bax KO mice were kindly provided by Dr. Woong Sun from the Department of Anatomy at Korea University College of Medicine. For generation of Bax KO bone marrow chimeric mice, C57BL/6 mice were sublethally irradiated with a dose of 8 Gy using the Theratron 780 system (AECL, Ottawa, Canada) and injected intravenously with 1 × 107 bone marrow cells from Bax KO mice. Reconstitution efficacy was assessed by genotyping leukocytes from peripheral blood obtained 3 weeks after bone marrow transplantation. To induce IRI, mice were anesthetized with the intraperitoneal (i.p.) injection of 15 mg/kg of ketamine (JHP Pharmaceuticals, LLC, Rochester, MI, USA) and 2.5 mg/kg of xylazine (Akorn Animal Health, Inc., Lake Forest, IL, USA) and subjected to bilateral renal pedicle clamping for 30 minutes via flank incision.
Drug administration
To test the effects of JAM-C blockade in vivo, a monoclonal anti-mouse JAM-C antibody (clone H33; Millipore, Billerica, MA, USA) or control immunoglobulin G (IgG) was administered i.p. at 1.5 mg/kg on days 1 and 2 following IRI induction.
Biochemical analysis
Three hundred microliters of blood were collected at 24, 72, or 120 hours after IRI induction. And serum creatinine was measured using a Hitachi 747 automatic analyzer (Black Scientific Inc., Bohemia, NY, USA).
Histologic examination
In periodic acid-Schiff (PAS) stain, histologic changes in the corticomedullary junction and outer medulla were evaluated semi-quantitatively, and the degree of tubular cell damage was graded on a scale of 1 to 4, according to the percentages per high-power field (HPF). The mean scores were compared between the groups. To detect neutrophil infiltration in the kidney, immunohistochemical staining with anti-mouse Gr-1 was performed. Interstitial fibrosis was assessed by Masson's trichrome (MT) staining. For quantitative analysis, ten random non-overlapping fields from the cortex to corticomedullary region were captured using an Olympus BX51 microscope and a 40× magnification lens (Olympus, Tokyo, Japan). The positive staining in the MT-stained sections was selected and semiquantitatively analyzed using the Leica Application Suite version 4.0 (Leica Microsystems, Heerbrugg, Switzerland).
Quantitation of cytokines/chemokines by cytometric bead array (CBA)
Quantitation of various cytokines and chemokines in the kidney tissue was conducted using a CBA, a bead based flow cytometric multiplexed immunoassay for simultaneous detection of multiple cytokines. A mouse inflammation kit (BD Bioscience, San Jose, CA, USA) was used to detect mouse interleukin (IL)-12p70, tumor necrosis factor (TNF)-α, interferon (IFN)-γ, monocyte chemotactic protein (MCP)-1, IL-10, and IL-6 from total kidney extracts. The mean cytokine expression level normalized to grams of protein in each group was expressed as the mean ± standard error (SE) and compared between groups.
Flow cytometry
Kidneys were minced and then treated with collagenase type IV (10 μg/mL; Sigma-Aldrich, St. Louis, MO, USA) and DNAse (Sigma-Aldrich). The digested kidney suspension was filtered through a 100-μm cell strainer (BD Bioscience), passed through a cotton column treated with 10% fetal bovine serum, and centrifuged at 3,000 rpm for 5 minutes. Antibodies were purchased from eBioscience (San Diego, CA, USA; CD45, Ly6G/Gr-1) and BD Bioscience (AAD and Annexin V). To detect ICAM-1+ neutrophils that remigrated from the kidneys into circulation, cells from peripheral blood were stained with anti-mouse Ly6G/Gr-1, anti-mouse intercellular adhesion molecule-1 (ICAM-1+; eBioscience), or anti-mouse Tamm-Horsfall protein (THP; BIOSS Antibodies, Woburn, MA, USA). Four-color fluorescence flow cytometric analyses were performed (FACSCalibur™; BD Bioscience) to determine the percentages of apoptotic neutrophils in the kidneys and ICAM-1+ reverse transmigrated neutrophils in peripheral blood. These data were analyzed using FlowJo software (Tree Star Inc., Ashland, CA, USA).
In vitro characterization of immune cells in Bax KO mice
To compare phenotype of immune cells, splenocytes from C56BL/6 or Bax KO mice were isolated. Ten million splenocytes were stimulated with 1 mM H2O2 for 6 hours, and the percentages of cells undergoing apoptosis (Annexin V+ 7-AAD−) were compared. To determine the innate immune function, 1 × 107 splenocytes were stimulated with 1 mM lipopolysaccharide (LPS; Sigma Aldrich Korea, Seoul, Korea) for 4 hours, and the production of TNF-α, IL-6, and IL-10 was compared. The migration of splenocytes toward the chemokine MCP-1 was evaluated using a transwell migration assay and the number of splenocytes migrated through porous membrane toward MCP-1 (1 mM) in the other side of transwell was compared.
Detection of THP in peripheral blood neutrophils
Peripheral blood mononuclear cells from sham-treated mice or mice 2 days after IRI were isolated and the level of THP was determined by enzyme-linked immunosorbent assay (ELISA; USCN Life Science, Houston, TX, USA) according to manufacturer instructions.
RESULTS
Creation of Bax KO bone marrow chimeras and characterization of Bax KO immune cells
Generation of Bax KOBM → wild-type (WT) chimera was confirmed by genomic DNA analysis of blood leukocytes. Absence of Bax gene was verified as previously described (4).
Bax KO splenocytes show partial resistance to H2O2-induced apoptosis compared to WT splenocytes, as expected (Fig. 1A). However, LPS-induced cytokine production and migration toward MCP-1 in Bax KO splenocytes are not different from those of WT splenocytes (Fig. 1B and 1C).
Fig. 1
Characteristics of Bax KO splenocytes. (A) Bax KO splenocytes are partially resistant to H2O2 induced apoptosis compared to WT splenocytes. (B) Cytokine production upon LPS stimulation was not different between Bax KO and WT splenocytes. (C) Migratory capacity of splenocytes toward MCP-1 was not different between Bax KO and WT. All experiments were conducted in triplicates.
KO = knockout, WT = wild-type, LPS = lipopolysaccharide, MCP = monocyte chemotactic protein, TNF = tumor necrosis factor, IL = interleukin, PBS = phosphate-buffered saline, HPF = high-power field.
*P < 0.05.

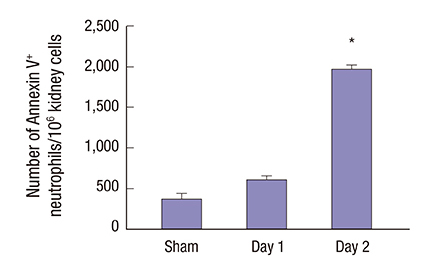
Apoptotic neutrophils identified during the recovery phase in the kidneys
While the kidney showed massive infiltration of live neutrophils one day 1 after IRI, the number of Annexin V+ Gr-1+ apoptotic neutrophils in the kidneys significantly increased on day 2 post-IRI. This suggests that some neutrophils began to die by apoptosis during the recovery phase (Fig. 2).

Clearance of neutrophils and functional, histological recovery in Bax KOBM → WT chimera
While infiltrated neutrophils almost disappeared by day 5 after IRI in WTBM → WT control mice, clearance of neutrophil is delayed (Fig. 3A) and tissue level of TNF-α, MCP-1, and IL-6 are persistently elevated in Bax KOBM → WT chimera (Fig. 3B). In addition, functional and histological recovery are significantly delayed in Bax KOBM → WT chimera in which only bone marrow derived cells are partially resistant to apoptosis (Fig. 3C and 3D).
Fig. 3
Neutrophil infiltration persisted in Bax KOBM → WT chimera throughout the recovery phase of IRI and was associated with delayed functional, histological recovery. (A) Left: representative pictures of Gr-1 immunohistochemistry, × 100, Right: mean number of Gr-1+ neutrophils in kidneys on day 1, 3, 5 of IRI. (B) TNF-α, MCP-1, and IL-6 levels in the kidneys were persistently elevated on day 5 of IRI in Bax KOBM → WT chimera. (C) Serum creatinine. (D) Left: representative pictures of PAS stained kidney tissues, × 100, Right: semiquantitative scoring of tubular injury (n = 5–6 per group).
KO = knockout, WT = wild-type, IRI = ischemia/reperfusion injury, TNF = tumor necrosis factor, MCP = monocyte chemotactic protein, IL = interleukin, WT = wild-type, PAS = periodic acid-Schiff.
*P < 0.05.
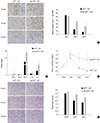
Persistent inflammation in Bax KOBM → WT chimera and fibrosis
Compared to WTBM → WT control mice, substantial fibrosis developed in Bax KOBM → WT chimera mice at 2 weeks after IRI (Fig. 4A). Interestingly, F4/80+ macrophages were predominant in the kidneys, and the number of macrophages significantly increased in Bax KOBM → WT chimera compared to control mice (Fig. 4B).
Fig. 4
Persistent inflammation in Bax KOBM → WT chimera led to early fibrosis. (A) Upper: representative pictures of MT stained kidney tissues, × 40, lower: semiquantitative scoring of fibrosis. (B) Upper: representative pictures of F4/80 immunohistochemistry, × 100, lower: mean number of F4/80+ cells in kidneys on 14 days after IRI (n = 5–6 per group).
KO = knockout, WT = wild-type, MT = Masson's trichrome, IRI = ischemia/reperfusion injury.
*P < 0.05.
Detection of circulating ICAM-1+ neutrophils during the recovery phase of IRI
In contrast to naïve murine neutrophils that do not express ICAM-1, neutrophils that have migrated twice in reverse direction from tissue through the endothelial barrier are known to express ICAM-1. By using flow cytometry, we observed an increase in ICAM-1+, Gr-1+ neutrophils in circulation on day 2 after IRI compared to those from day 1 after IRI, suggesting the possible occurrence of rTEM during the recovery phase (Fig. 5A and 5B).
Fig. 5
Circulating ICAM-1+ neutrophils increased during the recovery phase of IRI. (A) Flow cytometry showed the increase of ICMA-1+ Gr-1+ reverse transmigrated neutrophils in peripheral blood on day 2 after IRI. (B) The number of circulating ICMA-1+ Gr-1+ neutrophils out of 106 kidney cells (n = 4–5 per group).
ICAM-1 = intercellular adhesion molecule-1, IRI = ischemia/reperfusion injury.
*P < 0.05 compared to sham. †P < 0.05 compared to I/R day 1.

Identification of THP in circulating ICAM-1+ neutrophils
To confirm the origin of circulating ICAM-1+ neutrophils, we measured THP+ ICAM-1+ neutrophils in peripheral blood by flow cytometry. We first gated Ly6G/Gr-1+ neutrophils and then assessed the percentage of ICAM-1+ THP+ cells. Percentage of THP+ ICAM-1+ neutrophils significantly increased on day 2 of IRI, suggesting that these cells were derived from injured kidneys (Fig. 6A and 6B).
Fig. 6
Substantial portion of circulating ICAM-1+ neutrophils during the recovery phase of IRI are positive for THP. (A) Flow cytometry show the increased percentage of THP+ ICAM-1+ neutrophils on day 2 after IRI. Neutrophils were gated first. (B) Percentage of THP+ ICAM-1+ neutrophils after IRI. (C) THP was increased in peripheral blood leukocytes during the recovery phase of IRI (n=4–5 per group).
ICAM-1 = intercellular adhesion molecule-1, IRI = ischemia/reperfusion injury, THP = Tamm-Horsfall protein.
*P < 0.05.

We also isolated circulating leukocytes and measured the protein levels of THP. Compared to sham or day 1, THP levels in the leukocytes obtained from mice 2 days after IRI were significantly increased (Fig. 6C).
In vivo blockade of JAM-C and rTEM, recovery
Treatment with JAM-C blocking antibody starting 1 day after IRI resulted in increased circulating ICAM-1+ Gr-1+ neutrophils (Fig. 7A), suggesting that blocking JAM-C facilitated rTEM. This increase in ICAM-1+ Gr-1+ circulating neutrophils was associated with a significant increase in neutrophil clearance in the kidneys (Fig. 7B) and faster functional and histological recovery (Fig. 7C and 7D).
Fig. 7
Effect of JAM-C blocking antibody on the number of circulating ICAM-1+ neutrophils, kidney neutrophils, functional, and histological recovery following IRI. (A) Upper: Flow cytometry show that treatment with anti-mouse JAM-C blocking Ab resulted in increase of circulating ICAM-1+ neutrophils following IRI. Lower: percentage of circulating ICMA-1+ Gr-1+ neutrophils out of total neutrophils (n = 4–5 per group). (B) Treatment with anti-mouse JAM-C blocking Ab resulted in decreased number of infiltrating neutrophils on day 3 after IRI. Upper: representative pictures of Gr-1 immunohistochemistry, × 100, lower: mean number of Gr-1+ neutrophils in kidneys on day 3 of IRI (n = 4–5 per group). (C) Treatment with anti-mouse JAM-C blocking Ab resulted in faster functional recovery. (D) Treatment with anti-mouse JAM-C blocking Ab resulted in faster histological recovery. Upper: representative pictures of PAS stained kidney tissues, × 100, lower: semiquantitative scoring of tubular injury (n = 5–6 per group).
JAM-C = junctional adhesion molecule-C, ICAM-1 = intercellular adhesion molecule-1, IRI = ischemia/reperfusion injury, Ab = antibody, PAS = periodic acid-Schiff.
*P < 0.05 compared to day 1. †
P < 0.05 compared to day 2. ‡
P < 0.05.
DISCUSSION
In this study, we demonstrated that apoptosis and rTEM are critically involved in the clearance mechanism of neutrophil and recovery process following IRI. IRI and nephrotoxic insult to the kidneys provoke an inflammatory response contributing to tissue injury and dysfunction (1). Although the detrimental effects of acute kidney injury (AKI) on both short-term and long-term outcomes are well known, AKI is still a potentially reversible condition, suggesting the existence of natural processes that eliminate inflammation (56). To resolve inflammation, infiltration of inflammatory cells must be counterbalanced with effective clearance. This is especially relevant to neutrophils in kidney IRI because neutrophils are the most abundant leukocyte subset early in reperfusion period. In addition, recent study demonstrated somewhat distinct fate of infiltrated macrophages, another important leukocyte subset, to be a shift of their phenotype from proinflammatory M1 to anti-inflammatory M2 instead of being cleared (7).
Until recently, the resolution of inflammation has been considered a passive process. However, emerging evidence suggests that resolution is an active, tightly regulated process that induces the decline of proinflammatory mediators and the removal of inflammatory cells, leading to the restoration of normal structure (89).
The primary mechanisms of clearance of neutrophils are believed to be apoptosis. However, this speculation mainly comes from the in vitro observations that neutrophils undergo spontaneous apoptosis in standard culture conditions with a half-life shorter than 8 hours (1011). Few studies have demonstrated firm evidence that tissue-infiltrating neutrophils undergo apoptosis or that apoptosis is important as clearance mechanism and recovery in vivo.
In our study, we first observed an increase in the number of Annexin V+ apoptotic neutrophils on day 2 after IRI, whereas day 1 kidney was characterized by a huge infiltration of Annexin V− live neutrophils, confirming the occurrence of neutrophil apoptosis during the recovery phase. The critical role of apoptosis in the clearance mechanism or in recovery process was assessed in Bax KO bone marrow chimera, in which only the bone marrow-derived cells are partially resistant to apoptosis. This unique model system is thought to be a reasonable approach to examine the role of neutrophil apoptosis in vivo because of the possibility that Bax deficiency in tubular cell might have differential effect on kidney injury. Recent observation by Wei et al. (11) showing that only conditional Bax deletion in proximal tubules but not global Bax deletion ameliorated ischemic AKI support the differential effect of Bax deletion in proximal tubules or other cell types including leukocytes on IRI. The requirement of Bax gene in neutrophil apoptosis has been previously demonstrated (1213).
Although the number in outer medulla on day 1 after IRI were comparable, neutrophils persisted during the recovery phase in Bax KOBM → WT chimera compared to those in WTBM → WT control mice, in which neutrophils disappeared. In addition, the observation that higher tissue levels of proinflammatory cytokines associated with delayed functional and histological recovery in Bax KOBM → WT chimera could indicate the importance of neutrophil apoptosis in resolution of inflammation as well as the recovery process. These data are in concordance with several recent reports; Hanses et al. (14) demonstrated that in mice challenged with i.p. Staphylococcus aureus, reduced apoptosis of neutrophils in non-obese diabetic (NOD) mice compared to those from normolycemic NOD mice, led to reduced neutrophil clearance. Loynes et al. (15) used a transgenic zebrafish larva model, in which green fluorescent protein (GFP)-expressing neutrophils can be tracked in vivo, and demonstrated that agents that delay neutrophil apoptosis, such as LPS or caspase inhibitors, led to delay in the resolution of inflammation. By introducing a unique chimeric mouse model, our study convincingly demonstrates the critical contribution of neutrophil apoptosis to the natural clearance mechanisms of neutrophils and recovery process following IRI. Although we focused on neutrophils in this study, there is possibility that Bax KO chimera exert same effect on other leukocyte subsets. However, there were not apoptotic macrophages during the recovery phase (data not shown).
Apoptotic neutrophils are thought to be rapidly digested by macrophages, and thus, impairment not only in neutrophil apoptosis but also in any of the subsequent processes, such as in macrophage recognition or phagocytosis might lead to failure of the resolution of inflammation. Our lab previously showed that delayed macrophage depletion in the recovery phase was associated with persistent neutrophilic inflammation on day 7 after IRI, suggesting the important role of macrophages in resolution of inflammation (16). In addition, we also found that delayed neutrophil clearance in our system resulted in accelerated fibrosis on day 14 after IRI, indicating that the failure of resolution could lead to the development of chronic inflammation or fibrosis. However, there is still possibility that significantly higher number of macrophages in Bax KOBM → WT chimera is due to inhibition of macrophage apoptosis in fibrosis.
Until recently, cell death by apoptosis was thought to be the only fate of neutrophils that completed their effector function. However, the relatively small number of apoptotic neutrophils in this study led us to hypothesize that other mechanisms might also be involved in the clearance of neutrophils. Accumulating evidence suggests that neutrophils do not die immediately in tissue, but they can persist in the host by a process of reverse chemotaxis, known as rTEM. Typically, neutrophils are recruited to sites of infection or injury in a luminal to abluminal direction. However, neutrophils have been recently shown to undergo abluminal to luminal migration. Mathias et al. (17) observed that robust neutrophil chemotaxis to sites of wounding is followed by retrograde chemotaxis back into circulation in zMPO; GFP transgenic zebrafish by using in vivo time-lapse imaging, and suggested that this can be an important mechanism of resolution of inflammation. Therefore, on the assumption that neutrophils can make a U-turn into circulation, we hypothesized that rTEM could be a mechanism of resolution of inflammation in kidney IRI. At the same time, we asked how we could recognize those neutrophils that had undergone rTEM from injured kidneys. According to Buckley et al. (18), who developed a novel in vitro system that could recognize reverse-transmigrated neutrophils, neutrophils that underwent rTEM show distinct phenotypes: higher ICAM-1 expression, lower CXCR1 expression. Therefore, in our study, we first observed that the percentage of circulating ICAM-1+ neutrophils following IRI. ICAM-1+ reverse migrated neutrophils increased on day 2, while the most of circulating neutrophils on day 1 after IRI were ICAM-1−. Although this finding showed the increase of reverse-transmigrated neutrophils, it did not indicate that these cells were derived from the kidneys. Therefore, we tested whether these ICAM-1+ neutrophils contain THP, a kidney specific protein, and found that substantial portion of ICAM-1+ neutrophils on day 2 after IRI was also positive for THP. Although precise mechanisms of THP uptake are not clear, presence of THP in circulating neutrophils could suggest that these ICAM-1+ neutrophils were derived from the kidneys. Higher levels of THP in the leukocyte fraction obtained on day 2 after IRI compared to that on day 1 strengthens the hypothesis that some neutrophils infiltrated in the kidney after IRI made a U-turn back into circulation. The presence of kidney-specific THP in circulation following IRI was also recently demonstrated. El-Achkar et al. (19) showed that THP translocates from the apical to basolateral domain of the thick ascending limb, subsequently leak into the interstitium and vascular space during the recovery phase. They also showed a significant increase of serum THP on day 2 after IRI compared to baseline or day 1 levels, and ascribed this to the unique translocation of this protein into the interstitium. However, we noted that an increase in THP was more evident in the leukocyte fraction, and it is likely that THP from the interstitium is captured by some neutrophils and eventually appears in circulation possibly due to a process of rTEM.
Despite clear evidence indicating the occurrence of rTEM, little is known about the underlying mechanisms. Recently, Woodfin et al. (3) demonstrated that rTEM was associated with the loss of endothelial JAM-C. By using intravital confocal microscopy, they further showed that in vivo blockade or genetic deletion of JAM-C enhanced rTEM, suggesting that JAM-C functions as a negative regulator of rTEM. Therefore, we also tested the effect of in vivo blockade of JAM-C. Treatment with a JAM-C blocking antibody was initiated on day 1 after IRI to restrict the effect of JAM-C blockade to the recovery phase. Blocking JAM-C resulted in a significant increase in the number of circulating ICAM-1+ neutrophils and was also associated not only with increased clearance of kidney infiltrating neutrophils but also with faster functional and histological recovery. These results strongly suggest that rTEM of neutrophils is mediated by loss of JAM-C and contributes to the resolution mechanisms of neutrophilic inflammation as well as recovery process following IRI. Bradfield et al. (20) also observed a similar phenomenon and suggested that JAM-C forms an endothelial barrier that confines recently recruited cells to inflamed sites. Blocking JAM-C has also been demonstrated to facilitate recovery and clearance of neutrophil in cisplatin induced AKI (21). Although we did not track neutrophils directly, these data could suggest the important role of neutrophil rTEM in regulating the resolution of inflammation and recovery in kidney IRI.
In conclusion, we demonstrated that apoptosis and rTEM are critically involved in clearance of neutrophils in IRI. Although AKI frequently leads to chronic kidney disease (CKD), it is still reversible condition. Although the initial activation of the innate immune system is detrimental, there is an active, well-coordinated process for the resolution of inflammation, and this seems to be prerequisite for restoring normal function and structure. In clinical practice, the presence of CKD, repeated ischemia from intradialytic hypotension or concomitant use of nephrotoxicants might adversely affect this well-orchestrated resolution process leading to failure of recovery and progression to CKD. Better identification of pathways involved in resolution mechanisms as well as recognizing the pro-resolution properties of various drugs might help identify new therapeutic strategies to facilitate recovery from AKI.
 XML Download
XML Download