

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The bursa of Fabricius (BF) is a central humoral immune organ that is unique to birds and is equivalent to bone marrow in mammals. The BF contains a variety of bioactive substances, and bursin, the first peptide isolated from BF, selectively stimulated the differentiation of avian B lymphocytes precursor in vitro [35]. In recent years, various peptides have been isolated from BF, most of which have immunoregulatory functions [7891011172023]. In addition to their immune regulatory function, bursal peptides (BPs) derived from BF have exhibited other biological functions. For example, bursal septpeptides (BSP-I and BSP-II) induced both humoral and cellular immune responses in vivo [78]. Bursopentine (BP5), a newly isolated multifunctional active peptide, enhanced immune function in humoral and cellular immunity and had antioxidant function and antitumor activity [161718]. Given the relationships between peptides and various functions, it is vital to study BPs. In this study, four peptides (BP-I, BP-II, BP-III, and BP-IV) were isolated from BF and identified through RP-HPLC and MADIL-TOF methods [2122]. Previous study suggested that these peptides have the ability to promote B cell differentiation and have an immunomodulatory function on antigen-specific immune responses in chickens and mice. In this study, the potential immunoadjuvant properties of these four BPs were evaluated by detecting the humoral immunity and cell-mediated immune responses in mice coimmunized with BPs I to IV and an oil-formulated H9N2 avian influenza virus (AIV) vaccine. Viral titers and histopathological changes were assayed to assess the immunoprotective effect of BPs coinjected with H9N2 AIV vaccine against H9N2 AIV in mouse lungs.
Materials and Methods
Viruses, reagents, and peptides
The A/Chicken/Jiangsu/JS-1/2002 (H9N2) AIV was isolated and maintained in our laboratory [31]. Oil-formulated inactivated H9N2 AIV vaccine was purchased from Qian Yuan Hao (China). Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG was obtained from Boshide Corporation (China), and HRP-conjugated goat anti-mouse IgG1 and anti-mouse IgG2a were obtained from Bethyl Corporation (Germany). Fluorescein isothiocyanate-labeled anti-mouse CD3 and phycoerythrin-labeled anti-mouse CD4, CD8 antibodies were obtained from Caltag (China). Enzyme-linked immunosorbent assay (ELISA) cytokine kits were purchased from eBioscience (USA). RPMI 1640 medium was purchased from Procell (China). The four BPs (I–IV) were synthesized by Shanghai Science Peptide Biological Technology (China), and their purities were over 95%.
Vaccination immunization
The specific-pathogen-free (SPF) BALB/c mice (age, 4–6 weeks; body weight, 20 ± 2 g) were purchased from Henan Province Experimental Animal Research Center (China). The mice were divided randomly into seven groups of 30 mice each, half male and half female. On days 0 and 14, mice were intramuscularly immunized with either (i) 0.2 mL phosphate-buffered saline (PBS) as a negative control, (ii) 0.2 mL H9N2 AIV vaccine (107 TCID50; TCID50, 50% tissue culture infective dose), (iii) a mixture of 0.2 mL H9N2 AIV vaccine (107 TCID50) and BP-I (10 µg), (iv) a mixture of 0.2 mL H9N2 AIV vaccine (107 TCID50) and BP-II (10 µg), (v) a mixture of 0.2 mL H9N2 AIV vaccine (107 TCID50) and BP-III (10 µg), (vi) a mixture of 0.2 mL H9N2 AIV vaccine (107 TCID50) and BP-IV (10 µg), or (vii) not immunized or challenged and used as a blank control (Table 1). Details of the experimental time points are displayed in Fig. 1. This animal experiment was approved by the Institutional Animal Care and Use Committee of Henan University of Science and Technology (20170301001). All animals were humanely handled.
Hemagglutination inhibition assay
On days 0, 7, 14, and 21 after the first immunization, five mice from each group were randomly selected and blood serum samples were collected. Hemagglutination inhibition (HI) antibody titers were detected using standard HI microtiter assays [4].
Specific antihemagglutinin IgG antibody assay
On days 7 and 21 after the first immunization, specific antihemagglutinin (anti-HA) IgG antibody titers of serum were analyzed by ELISA [32]. Briefly, ELISA plates were coated with a purified prokaryote-expressed recombinant HA protein (expressed in Escherichia coli BL21, 10 µg/mL) [35]. Aliquots of serum were added to the plates, incubated overnight, washed, and then incubated with HRP conjugates of goat anti-mouse IgG, IgG1, and IgG2a. Finally, tetramethylbenzidine substrate was added and the reaction was stopped. Titers at half maximal optical density were determined by linear interpolation. Each serum sample was assayed in triplicate.
AIV-neutralizing antibody determination
Serum samples collected from each group of mice on 21 days after first immunization were incubated with 100 plaque-forming units (PFU) of H9N2 AIV A/chicken/Jiangsu/JS-1/2002, and the titers of the H9N2 AIV-neutralizing antibody were determined as described previously [27].
Cytokine assays
On days 7 and 21 after the first immunization, the levels of interleukin-4 (IL-4) and interferon gamma (IFN-γ) in mouse serum were detected using the ELISA method, according to the ELISA cytokine kits' instructions.
Detection of cytotoxic T-lymphocyte assay
To investigate the effect of BPs I to IV on CTL responses, mouse sera on day 21 after the first immunization were analyzed according to the instructions for the CytoTox 96 Non-Radioactive Cytotoxicity Assay kits (Promega, USA). Cytotoxic assays for effector cells were performed as described previously [34]. NIH3T3 cells, used as target cells, were infected with AIV A/chicken/Jiangsu/JS-1/2002 (H9N2) (moi 20 PFU/cell) for 24 h [13]. The specific methods of operation and calculating the specific killing percentage are as described previously [19].
Spleen T-cell subtyping
Spleen lymphocytes were isolated in samples on day 7 after the second immunization. Flow cytometric analysis was used to analyze the expression of spleen T-cell CD3+ and their subtypes CD3+CD4+ and CD3+CD8+ [32].
Virus challenge experiment
Two weeks after the second immunization, 15 mice per group were perorally challenged with 2.5 × 106 TCID50 AIV A/chicken/Jiangsu/JS-1/2002 (H9N2) plus 0.2 mL PBS. Lung samples from 5 mice per group were collected at 3, 5, and 7 days after virus challenge and stored. Viral copies in the lungs of the mice were determined by using real-time polymerase chain reaction (RT-PCR). The PCR primers were designed based on the HA gene sequence of avian influenza H9N2 virus strain JS-1 (A/Chicken/Jiangsu/JS-1/2002) in GenBank (accession No. AY364228). Amplification was performed using SYBR green according to a previously described method [24]. The standard curve for RT-PCR quantification was constructed using the HA gene in vector pET32a-HA (H9N2), which was kindly provided by Qisheng Zheng (Institute of Veterinary Science, Jiangsu Academy of Agricultural Sciences, China). Virus titers in infected mouse lungs were determined by using a TCID50 assay [29]. Briefly, mouse lung was homogenized in virus growth medium (VGM) (10% wt/vol) in which the Dulbecco's modified Eagle's medium contained an antibiotic-antimycotic (HyClone; Thermo Scientific, USA) as well as 1% bovine serum albumin. Ten-fold serial dilutions of the sample were added in quadruplicate to Madin-Darby canine kidney cells that had been seeded in microwell plates 1 day earlier and allowed to absorb for 2 h at 37℃ in an incubator. Fresh VGM was then added to the cells, and the cells were incubated at 37℃ for another 48 h. Then, the culture supernatants were mixed with the same volume of 1% (vol/vol) SPF chicken red blood cells (in PBS) and incubated for 15 to 20 min at room temperature. The virus titers were calculated and expressed as the log10 TCID50 per milliliter of lung tissue.
Histology
Mouse lung tissues were dissected at 3, 5, and 7 days post-challenge and gently fixed in buffered formaldehyde solution, dehydrated in a graded series of ethanol, and embedded in paraffin. Sections were cut and stained with H&E [26]. Histopathological changes of the lung section were scored as described previously [29].
Statistical analysis
All statistical analyses were performed by using GraphPad Prism 6 software (GraphPad Software, USA). Differences were considered significant at p < 0.05 or p < 0.01. Statistical analyses were performed using unpaired t-tests or one-way ANOVA F-statistics. Data are presented as mean ± SD values. Differences among the six experimental groups (five immunized and one control group) were assessed by using Tukey's multiple comparison tests.
Results
BPs and antigen-specific immune responses
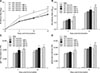
To evaluate antigen-specific immune responses, mice coimmunized with inactivated H9N2 AIV and BPs, the levels of HI antibody, anti-HA antibody, and IgG (IgG1, IgG2a) antibody titers were examined. Assays of serum HI antibody titers 0, 7, 14, and 21 days after initial immunization were observed (panel A in Fig. 2). As shown, mice coimmunized with H9N2 AIV vaccine and BP-I or BP-III induced HI antibody responses, though the changes were not significant (p > 0.05). Mice coimmunized with H9N2 AIV vaccine and BP-II produced significantly high HI antibody titers on day 21 (p < 0.05). Mice coimmunized with H9N2 AIV vaccine plus BP-IV produced significantly high serum HI antibody titers at days 7, 14, and 21 after the first immunization (days 7 and 14, p < 0.05; day 21, p < 0.01). As shown in panel B in Fig. 2, compared to mice immunized with the H9N2 AIV vaccine alone, mice coimmunized with H9N2 vaccine plus BP-II produced high HA antibody titers at day 21 post-immunization (p < 0.05). Mice coimmunized with H9N2 vaccine plus BP-IV induced an even higher HA antibody titer at days 7 and 21 post-immunization (p < 0.05 and p < 0.01, respectively). No significant induction of anti-HA antibody titers was observed in mice coimmunized with the H9N2 AIV vaccine plus BP-I or BP-III (p > 0.05). The IgG subtype assay results (panels C and D in Fig. 2) showed that the BP-II group induced IgG1 and IgG2a production at 21 days post-immunization (p < 0.05). Moreover, the BP-IV group induced both IgG1 (days 7 and 21, p < 0.05) and IgG2a (day 7, p < 0.05; day 21, p < 0.01) antibody responses. In contrast, IgG (IgG1, IgG2a) antibody titers were not induced significantly in mice coimmunized with H9N2 AIV vaccine plus BP-I or BP-III (p > 0.05).
It is known that the production levels of neutralizing antibodies and antibodies are not always consistent. Therefore, we tested whether BPs can increase the production of virus neutralizing antibodies. The results showed that the levels of neutralizing antibody titers were in accordance with the antibody titer levels; neutralizing antibody titers in mice coimmunized with BP-II or BP-IV plus H9N2 AIV vaccine were significantly higher than those in mice immunized with the H9N2 AIV vaccine alone (days 7 and 21, BP-II, p < 0.05; days 7 and 21, BP-IV, p < 0.01) (Table 2). No significant induction of antibody production was observed in the BP-I and BP-III groups (p > 0.05). The results indicated that BP-II and BP-IV significantly stimulate the production of antigen-specific antibody titers.
BPs and the production of Th1- and Th2-type cytokines
The levels of Th1 (IFN-γ)- and Th2 (IL-4)-type cytokines in mice coimmunized with the inactivated H9N2 AIV vaccine and BPs were also determined. Serum cytokine concentrations were assayed 7 and 21 days after the first immunization (panels A and B in Fig. 3). On days 7 and 21 post-immunization, mice coimmunized with H9N2 AIV vaccine and BP-I or BP-II produced a significant increase in IL-4 cytokine level (BP-I, p < 0.05; BP-II, p < 0.05) compared to that in mice in the vaccine control group. Additionally, mice coimmunized with H9N2 AIV vaccine and BP-I or BP-II showed a significant increase in IFN-γ cytokine level on days 7 and 21 (BP-I, p < 0.05; BP-II, p < 0.05). However, no significant production of IL-4 and IFN-γ cytokines were observed in the BP-III group (p > 0.05). Mice coimmunized with H9N2 AIV vaccine and BP-IV showed significant increases in the levels of both IL-4 (days 7 and 21, p < 0.01) and IFN-γ cytokines (day 7, p < 0.05; day 21, p < 0.01).
BPs on specific CTL responses
By performing released lactate dehydrogenase (LDH) cytotoxicity assays, we observed that mice coimmunized with H9N2 AIV vaccine and BP-I or BP-IV had higher levels of specific CTL responses to H9N2 virus than that in mice immunized with H9N2 AIV vaccine alone (BP-I, p < 0.05; BP-IV, p < 0.01) (panel C in Fig. 3).
Changes in T-cell subsets
The percentages of total CD3+T cells and related subsets (CD4+T cells [CD3+CD4+] and CD8+T cells [CD3+CD8+]) in the splenic lymphocytes were assessed in mice coimmunized with a mixture of H9N2 AIV vaccine and BPs (panel D in Fig. 3). The results showed that the addition of BP-I and BP-II significantly increased the percentages of overall CD3+T cells (p < 0.05) and the CD4+T-cell subset (BP-I, p < 0.01; BP-II, p < 0.05). Notably, BP-I also significantly induced the percentage of the CD8+T-cell subset (p < 0.05). In addition, BP-IV significantly increased the percentages of not only the overall CD3+T cells (p < 0.01) but also those of both the CD3+ CD4+ (p < 0.01) and CD3+CD8+ (p < 0.05) T cells.
Immunological protection against H9N2 AIV challenge
Mouse lung lobes were harvested at days 3, 5, and 7 after virus inoculation. Some of the lobes were used in the determination of viral loads by using RT quantitative PCR (RT-qPCR; panels A–C in Fig. 4) as well as in the determination of the TCID50 of the virus (panels D–F in Fig. 4). The RT-qPCR assay detected AIV copies in mouse lung samples (panels A–C in Fig. 4). As shown, numbers of lung viral copies were significantly reduced in the mice coimmunized with H9N2 AIV and BP-I or BP-II compared to mice immunized with H9N2 AIV vaccine alone on days 3, 5, and 7 post-immunization (p < 0.05). The number of lung viral copies was even more reduced in mice coimmunized with H9N2 AIV vaccine and BP-IV on day 7 (p < 0.01). Furthermore, mice coimmunized with H9N2 AIV vaccine and BP-I or BP-II exhibited virus removal from the lungs at 3 days after the challenge (p < 0.05). The group of mice coimmunized with H9N2 AIV vaccine and BP-IV had significantly reduced AIV viral titers in lungs on days 3 and 5 after challenge (p < 0.01). Moreover, there were almost no virus particles detected in the lungs of mice coimmunized with H9N2 AIV vaccine and BP-IV at day 7 after challenge (panel F in Fig. 4).
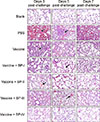
The remaining lung lobes harvested at days 3, 5, and 7 after viral infection were H&E stained (Fig. 5) and scored for pathology signs (Fig. 6). The lung tissues of mice in the PBS group were seriously damaged and exhibited severe perivascular and interstitial infiltrates. The pathology assessments of the lungs revealed that those in mice of the H9N2 AIV vaccine and BP-IV group were close to normal 7 days after infection (Fig. 5) with that group's pathology score lower than those of the H9N2 AIV vaccine group and the other H9N2 AIV vaccine and BP groups (panel C in Fig. 6). The BP-I and BP-II groups showed only slight infiltration and had lower pathology scores, whereas the H9N2 AIV vaccine and BP-III group showed severe perivascular infiltration with a higher pathology score than those of the BP-I and BP-II groups and showed similar pathology changes to those of the H9N2 AIV vaccine group (Fig. 5). Therefore, the pathology scores were in accordance with the lung viral titer results and the observed pathological changes in the mouse lungs in each group.
Discussion
Vaccines are essential for controlling the spread of infectious diseases in humans and animals. The appropriate adjuvant with the vaccine can extend the time of the specific response and effectively improve the immune response [25]. However, most adjuvants have side effects [61530]. Thus, it is essential to identify new and optimal adjuvant candidates. It has been reported that some small peptide immunostimulants can be used as vaccine adjuvants [132]. Liu first reported four new BPs from the avian immune system, and those four peptides have been reported to have immunomodulating activities and immunomodulatory functions in antigen-specific humoral and cellular immune responses [2122]. In this study, we investigated the adjuvant activity of BPs I to IV in mice challenged against the H9N2 AIV.
In this study, inoculation of a mixture of H9N2 AIV vaccine and BPs induced different levels of humoral and cell-mediated immune responses in mice. When coinjected with the commercial H9N2 AIV vaccine, BPs I to IV, being used as adjuvants, produced different titers of antibodies involved in humoral responses. Mice immunized with H9N2 AIV vaccine and BP-II exhibited high antibody titers, including HI antibody titers, antigen-specific HA antibody titers, as well as IgG1 and IgG2a production, especially AIV-neutralizing antibody titers in mice. Antibody titers in mice coimmunized with H9N2 AIV vaccine BP-I or BP-III produced no change from that in mice immunized with the H9N2 AIV vaccine alone. Furthermore, mice immunized with the H9N2 AIV vaccine and BP-IV produced more significant changes in the levels of HI, HA, and AIV-neutralizing antibody titers than those from coimmunized with the other three BPs. This result suggested that BP-II and BP-IV can enhance specific humoral immune responses when combined with the H9N2 AIV vaccine.
As has been reported, cellular immunity also has a key role in fighting virus infections [14]. In the humoral response assessment, we observed that BP-II and BP-IV induced both IgG1 and IgG2a antibody responses. The IgG1 and IgG2a antibody isotypes are indicators of Th1 and Th2-type immune responses, respectively, and Th1 and Th2 cytokines have important roles in the complex immune system [21228]. These results indicate that BP-II and BP-IV can induce a mixed Th1 and Th2 immune response. The effect of BPs on cell-mediated immunity was examined in vaccinated mice by performing cytokine assays. We determined the levels of Th1- (IFN-γ) and Th2- (IL-4) type cytokines in mice coimmunized with the inactivated H9N2 AIV vaccine and BPs. Interestingly, we observed that, in addition to BP-II and BP-IV, BP-I also increased IL-4 and IFN-γ cytokine secretion, thus affecting the CD4+T-cell subsets. CD4+T cells can be disvided into Th1- and Th2-type cells according to their cytokine secretion. Lymphocyte homeostasis is essential for immune system balance [33]. Thus, flow cytometric analysis was used to measure the immunophenotyping of T lymphocytes including mice peripheral blood CD3+T cells and their subset CD4+ (CD3+CD4+) T cells and CD8+ (CD3+CD8+) T cells. Peripheral blood T-cell subtypes results showed that BP-I, BP-II, and BP-IV significantly increased the percentage of overall CD3+T cells and subset CD4+T cells. In addition, BP-I and BP-IV significantly increased the percentage of subset CD8+T cells. Furthermore, we found that CTL responses were higher in mice coimmunized with H9N2 AIV vaccine and BP-I or BP-IV than that in mice immunized with the H9N2 AIV vaccine alone. CTLs are known as “killer” cells because they can kill target cells in response to presentation of a specific antigen. The major surface marker of CTLs is CD8+. Taken together, the results show that BP-I produces a better cellular immune response than BP-II and BP-III, while BP-IV has an effective adjuvant activity in vaccines that enhances humoral and cell-mediated immune responses.
To further evaluate the effect of BPs as adjuvants in immunity protection, mice were challenged intranasally with H9N2 AIV (A/Chicken/ Jiangsu/JS-1/2002) on day 28 after they had been coimmunized with H9N2 AIV vaccine and BPs. As the A/chicken/Jiangsu/JS-1/2002 H9N2 AIV is a low-pathogenicity AIV, and as all challenged mice survived the infections [16], we used RT-PCR as well as TCID50 to detect the challenge virus in the lungs on days 3, 5, and 7 post-infection. Three days after the challenge, BP-I, BP-II, and BP-IV inhibited replication of the virus in mouse lungs. After coimmunization with H9N2 AIV vaccine and BP-I or BP-II, virus titers were significantly lower in the lungs of the coimmunized mice than in mice immunized with the H9N2 AIV vaccine at both 3 and 5 days after the challenge. Moreover, there were almost no detectable virus particles in the lungs of mice immunized with BP-IV as the adjuvant at 7 days post-challenge. Compared to the other three peptides, mice vaccinated with BP-III as adjuvant had higher viral titers on days 3 and 5, which was similar to the titer levels in mice immunized with the H9N2 AIV vaccine. As viral copies and viral titers only calculate the number of viri and cannot directly reflect the status of lung lesions, we assessed the pathological changes and pathology scores in mouse lungs. The inflammasome responses in mouse lungs were reduced in the BP-I, BP-II, and BP-IV groups on days 3 and 5 after virus infection, with only slight infiltration observed in the BP-I and BP-II groups on day 7. The BP-IV group's condition was close to normal on day 7. Similar to the viral titers, BP-III showed no obvious protection of mouse lungs compared with that in the vaccine group. Our data show that the pathological changes and pathological scores were in accordance with lung virus titers, and the results confirm that BP-IV provides strong protection against H9N2 AIV infection, which indicates it has the potential to be used in vaccine formulations.
In summary, the immunoadjuvant activities of four BPs derived from BF were studied. On the whole, BP-I primarily induced cell-mediated immune responses. BP-II significantly induced humoral immune responses. BP-III did not have a significant effect on activity in humoral and cell-mediated immune responses compared with that of the control group. BP-IV extraordinarily enhanced both humoral and cell-mediated immunity, provided protection against H9N2 AIV virus infection in mouse lungs; thus, BP-IV has adjuvant activity and may be effective for adjuvant applications. That four peptides displayed different levels of humoral and cell-mediated immune responses suggesting that BPs contained in BF act as multifunctional peptides and can exert different immune functions in mice, indicating that the various small molecules contained in BF can have diverse roles; these differences may balance the immune microenvironment. Although BP-III did not display significant adjuvant activity, we speculate that further research on the immune activity of BP-III will contribute new insights into other functions.






 XML Download
XML Download