

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
With no exception in the field of allergy, cases of pollen/food-associated syndromes (PFAS) are increasing. Between 20% to 40% of pollen-sensitized patients are also sensitized to fruits worldwide, with peach, apple and kiwi in the top 3 fruits.1 Many efforts have been put on the study of the birch/apple syndrome,2 but few data are reported on the cypress/peach syndrome.3 Recently, we have shown that immunoglobulin E (IgE) cross reactivity between peach and Italian cypress (Cupressus sempervirens, Cups) pollen is borne by members of the Gibberellin-regulated protein (GRP) family present in both allergenic sources, BP14 in cypress pollen and Pru p 7 in peach.4 GRPs are small cationic proteins, expressed in plant in response to a biotic or abiotic stress.5 They exhibit anti-microbial properties,6 and 4 allergens from this protein family are characterized in fruit (peach, orange, pomegranate and Japanese apricot) and 1 in pollen (Cups pollen).
To assess the clinical relevance of cross-sensitization to BP14 and Pru p 7, the case of a patient suffering since childhood from cypress pollen allergy and also food allergy to fruits including peach and pomegranate was thoroughly explored clinically (anamnesis and skin prick test [SPT]) and biologically using not only conventional diagnostic techniques to study the serum IgE specificities (single and multi-arrays) but also immunoprints and a basophil activation test (BAT) with total allergenic sources extracts and purified allergens.
MATERIALS AND METHODS
Cypress pollen and fruit protein extractions
Cups pollen grains (10%, w:v) (Allergon AB, Ängelholm, Sweden) were incubated overnight in phosphate buffer saline (PBS) pH 7.4 at 4°C by rotation. After centrifugation (18,000 g, 4°C, 20 minutes), the supernatant was dialyzed against distilled water and lyophilized. The lyophilized extract was dissolved in distilled water and stored as aliquots at −20°C until use.
Peach, grapefruit, orange and pomegranate were purchased in market. The peach was prepared by crushing the fruit peel. The grapefruit and orange pulps were deprived of juice by pressing and the remaining insoluble part used for protein extraction. The pulp pomegranate was prepared after removal of the seed. These 4 preparations were then submitted to protein extraction using a multidimensional grinder FastPrep-24 (MP-Biomedicals, Illkirch-Graffenstaden, France), for 40 seconds at 4°C at 6 m/sec using matrix C (1 mm diameter silica beads) in presence of PBS. After centrifugation (18,000 g at 4°C, 20 minutes) supernatants were collected and stored as aliquots at −20°C until use.
Native and recombinant proteins
Recombinant snakin-1 and peamaclein
Recombinant snakin-1 and peamaclein were prepared in Pichia pastoris and purified essentially as described in Kuddus et al.6 Briefly, after induction phase, yeast pellet and supernatant were separated by centrifugation (14,000 g, 4°C, 30 minutes). The protein containing supernatant was purified on a cation exchange column followed by a RP-HPLC, lyophilized and stored at −30°C.
BP14 from cypress pollen
Pieces of gel containing BP14 were excised from a sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) 8%–18% where only cypress pollen basic proteins were run.7 The in-gel BP14 was then eluted from the gel with distilled water in a multidimensional grinder MP-Biomedicals FastPrep-24 for 40 seconds at 6 m/sec with 1 mm diameter silica beads. The beads, acrylamide residues and the BP14-protein were then filtered on an ultrafree-MC filter (5 µm; Millipore, Billerica, MA, USA). The filtrate containing BP14 was stored at −20°C. MS/MS experiments were performed on the purified protein which confirms that it is a GRP (data not shown).
Patients and their sera
The cypress pollen and food allergic patient was selected based on clinical symptoms (rhinitis, conjunctivitis and asthma) and SPT using cypress and birch pollen extracts. Sensitization was determined with either individual classical fluorescence enzyme immunoassay (FEIA) (Thermo Fischer Scientific, Villebon, France; Phadia, Uppsala, Sweden) on the ImmunoCAP 2500 apparatus, according to the manufacturer's recommendations with a detection limit set at 0.10 kU/L or using 2 types of IgE multiarray microchips: ISAC (Thermo Fischer Scientific) and FABER (CRDiagnostics SAS, Bullion, France; a partner of P-Friendly Allergen Nano-Bead Array) with a detection limit set at 0.30 ISU-E and 0.10 FIU/mL, respectively. Written informed consent was obtained from the patients.
The serum from a healthy individual (non allergic, non atopic) and sera from atopic patients with other types of allergy were used as negative controls and corresponded to residues from biological analysis laboratories for the diagnosis of allergy. The sera were stored at −20°C until use.
Gel electrophoresis analysis
Proteins were separated in unreduced conditions in 1-dimensional gel electrophoresis (1-DE) SDS-PAGE in a 8%–18% gradient polyacrylamide gel (ExcelGel; GE Healthcare, Uppsala, Sweden) on a flat-bed electrophoretic chamber (Multiphor II; GE Healthcare) at 12°C. Molecular mass (Mr) markers (GE Healthcare, Little Chalfont, UK) ranging from 14.4 to 94 kDa were used as comparative references. Separated proteins were then either electrotransferred onto a 0.2 µm cyanogen bromide activated nitrocellulose (NCa) membrane (Optitran®BA-S 83; Schleicher and Schuell, Dassel, Germany) for immunoblotting assays or stained with Coomassie blue or silver nitrate for protein detection.
Cups protein extract was submitted to double one-dimensional gel electrophoresis (D1-DE) with basic proteins (pI 8.5–9.5) previously separated in isoelectric focusing (pH 2–11) and to classical two-dimensional gel electrophoresis (2-DE) according to Shahali et al.7 Isoelectric point markers (Serva, Heidelberg, Germany) ranging from 4.45 to 9.6 were used as comparative references in first dimension.
Immunoblotting
Immunoblotting was performed as described in Shahali et al.7 Briefly, NCa membranes, either intact for 2-DE or cut in 2.5-mm wide strips for 1-DE and D1-DE screening, were first incubated with serum (1:10 dilution) followed by alkaline phosphatase-conjugated goat anti-human IgE Fcε-specific antibodies (Sigma-Aldrich, St. Louis, MO, USA) and then substrate. For each immunoblot analysis, no serum and the serum from a healthy individual (non atopic, non allergic) were used as negative controls.
BAT
Ethylenediaminetetraacetic acid anti-coagulated whole blood was mixed with a Research-Use-Only (RUO) dried down antibody panel (Duraclone) specific to CD45, CD3, CRTH2, CD203c and CD63 in addition to activation buffer (Beckman Coulter, Marseille, France) and considered allergenic sources, further incubated at 37°C for 15 minutes. Extracts and purified proteins were the same as those used for immunoblots. After incubation, red blood cells were lysed using CE-IVD OptiLyse C lysis solution and associated protocol (Beckman Coulter) and further analyzed on a RUO 13-color Cytoflex flow cytometer (Beckman Coulter, Miami, FL, USA). Basophils were gated as CD45+, CD3−, low SS and CRTH2+ cells. Activation level was characterized by monitoring the expression of CD203c and CD63. Positive control tubes correspond to anti-IgE stimulation and are used to verify the presence of ex vivo reactivity of the considered whole blood sample. The Duraclone based approach for BAT that has been used herein is for RUO and not for diagnostic procedures.
RESULTS
Clinical evaluation
The studied patient is a 40 year-old-man currently living in Paris (northern France) and born in South-west France. He experienced an anaphylactic shock after ingestion of pomegranate (Punica granatum, Lythraceae family) and strong oral syndrome after ingestion of Rosaceae fruits (apple and peach). He has seasonal rhinoconjunctivitis during the cypress and birch pollen seasons which overlap in the north of France, relieved by antihistaminic treatment. SPT are positive with birch and cypress pollen extracts.
Biological evaluation: serum IgE binding
Specific IgE antibodies to cypress and birch pollen, peach, citrus, apple, strawberry, kiwi and cherry were found using either an individual classical FEIA or 2 types of IgE multiarray microchips (Table). Despite some inconsistencies, the 3 assays were coherent for IgE reactivities to PR10 allergenic proteins from various sources and also for the major allergens from Cupressaceae pollen Cup a 1 and Cry j 1. No IgE binding was found against LTPs or profilins.
Table
Specific IgEs of studied specimens (case and control) against various extracts or allergens using either an individual classical fluorescence enzyme immunoassay (CAP, kUA/L) or two types of IgE multiarray microchips: ISAC (ISU-E) and FABER (FIU/mL)

Colors are standardized according to the proposed ISAC scale: red, very high; orange, moderate/high; yellow, weak; grey, very weak; and 0, undetectable.
IgE, immunoglobulin E.
![]()
After separation of proteins by 1-DE or D1-DE, IgE immunoblots were performed on proteins extracted from Cups pollen, potato and fruits reported to contain allergenic GRP: peach, citrus (orange and grapefruit) and pomegranate. Purified native or recombinant proteins from cypress pollen (BP14), peach (peamaclein, Pru p 7) and potato (snakin-1) were also used (Fig. 1A). Specific IgE antibodies against BP14, a 14 kDa cationic protein were evidenced in the total Cups extract (lane 1) and the basic protein fraction (lane 2), and was confirmed on the native purified BP14 (lane 3). The additional IgE reactivity at 43 kDa corresponds to the binding to the major allergen Cup s 1 and Cup s 28 (Fig. 1A, lanes 1 and 2; Fig. 1B). In peach, IgE reactivities are found against a protein of 20 kDa (lane 4), likely to be Pru p 1 and another of 14 kDa, probably Pru p 7, since Pru p 7 is recognized in purified recombinant form by the patient's serum IgE (lane 5). Moreover, an IgE binding was observed against a single 14-kDa protein in the pomegranate extract (lane 6) in agreement with the severe symptoms reported by the patient upon ingestion of pomegranate. Concerning citrus, only a very weak (non cationic, data not shown) IgE reactivity towards a 15-17 kDa protein was detected in orange and grapefruit pulp extracts (lanes 9 and 10). The recombinant snakin-1 (lane 8) reported to cross-react with BP14 and to share 82% sequence identity with Pru p 74 was recognized by the patient's IgE, and a similar reactivity was also found in total crude potato extract (lane 7). Immunoblot performed with 2-DE separated proteins from Cups pollen extract confirmed the IgE binding pattern to Cup s 1, Cup s 2 (cationic 43 and 55 kDa) and BP14 allergen (Fig. 1B).
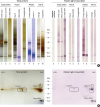 | Fig. 1IgE immunoreactivity to extracts and proteins. (A) SDS-PAGE coomassie blue (lanes 5, 7, 8) or silver nitrate (lanes 1, 2, 3, 4, 6, 9, 10) stained proteins. IgE immunoblots with patient serum (lanes 0 to 10). Negative controls (B) 2-DE. Left panel: silver-stained total proteins. Right panel: IgE immunoblot with patient serum. Relative molecular masses in kDa.SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis; 2-DE, two-dimensional gel electrophoresis.
|
Biological evaluation: BAT
Finally, the use of BAT allowed us to demonstrate that the basophils of this patient could be activated ex vivo with total protein extracts from cypress pollen, peach, pomegranate or orange and also with purified proteins such as BP14 and Pru p 7. Both activation markers, CD63 and CD203c, were overexpressed in a dose-dependent manner following addition of allergenic sources to basophils (Fig. 2). Interestingly, while IgE binding of patient's IgE to snakin-1 was observed (Fig. 1A), no basophil activation was shown, in keeping with the tolerance to potatoes mentioned by the patient. Considering the observed ex vivo activation of basophils from a control atopic patient, not allergic to cypress pollen but to birch and alder pollen and some foods (Fig. 2 and Table), the used BAT assay might be considered specific in the studied case. Indeed, the basophils from this control atopic patient were not activated ex vivo by cypress pollen, orange, pomegranate or snakin-1 and only weakly activated by BP14 and peach extracts, in agreement with the results of the presence of specific IgEs to peach (Table).
 | Fig. 2Basophil activation test: ex vivo activation of patient's basophils expressing CD203c (A) or CD63 (B) standardized activation markers (anti-IgE, 100% activation), (no allergen, 0% activation). Four serial concentrations were used: 10 to 104 ng/mL of each extract, 1 to 103 ng/mL for recombinant proteins and 5 to 5 × 103 ng/mL for native BP14. One concentration of each allergenic source was used for the control patient's basophils.
|
DISCUSSION
The results show that BP14, the only pollen allergen member of the GRP protein family reported so far, is able to induce ex vivo mediator release from basophils of a cypress allergic patient. It is therefore strongly suggested that this allergen is clinically relevant like the cross-reactive allergen Pru p 7 in peach and we can therefore propose that these ex vivo reactivities, associated with previously published cross-inhibitions,9 are on the basis of the reported PFAS cypress/peach described by Hugues et al.3 in 2006.
IgE reactivity was found against a 14 kDa protein in pomegranate extract otherwise able to activate patient's basophils. Despite the negative results obtained with pomegranate allergens tested on the microarray chip, it is tempting to speculate that the IgE binding protein in pomegranate is Pun g 7,10 because Pun g 7 shares structural (Mr, charge, member of GRP family) and functional properties (allergenicity) with BP14 and Pru p 7.11
GRPs are diversified in structure comprising a variable domain in size and amino acid sequence with a conserved C-terminal domain.5 Snakin-1 belongs to this protein family. It shares 82% sequence identity with Pru p 7 and was shown to cross-react with BP14,49 although to our knowledge, the snakin-1 from potato has not yet been described as an allergen. Moreover, in contrast to Pru p 7, snakin-1 was unable to inhibit the binding of specific IgE to BP14 and to activate patient's basophils which may be related to a lower affinity of cross-reactive BP14 specific IgE antibodies for snakin-1 than for Pru p 7.
Taken together, the results further gain insights into the properties of allergen members of this newly described GRP family. Whether other GRPs in pollen and/or food are specific markers for PFAS remain to be studied.
 XML Download
XML Download